
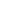


Hagin basoen historia Iberiar Penintsularen Iparraldean
17-12-2020
Denboran zehar lurrak eraldaketa ugari izan ditu planetako bizitza baldintzatu zutenak. Lurraren historiako alterazio nagusiak kontinenteen jitoa eta klima aldaketa izan ziren. Iraganean klima aldaketaren indar garrantzitsuena lurraren orbita-zikloen aldaketak ziren, azken ondorio modura planetako zati bakoitzak jasotzen duen intsolazio kopurua eta, horrenbestez, planetako zona ezberdinetako klima baldintzatzen dutenak. Haatik, azken urteotan lurraren historia gizakiak ekosistema naturaletan eragindako alteraziotik eratorritako aldaketak dira ezaugarri.
Informazio paleoekologikoak klimaren bilakaera eta bioma eta espezie ezberdinengan horrek duen eragina ezagutzeko aukera ematen digu. Horri dagokionez, landare arrastoen ikerketak ekosistemen eta espezieen dinamikaren inguruko ikuspegi zehatzenetako bat ematen digu. Azterketa paleobotanikoek espezie ezberdinek klima aldaketa edo asaldura ezberdinen aurrean duten erantzuna aztertzeko aukera ematen digute.
Zuhaitz espezieetako makrofosilen (enbor, adar, hosto edota fruituen aztarnak) eta mikrofosilen (polen-aleak) azterketatik abiatuz horiek lurralde jakin batean izandako bilakaera berreraiki daiteke. Hala, horietan izandako alterazio natural eta antropikoek denboran zeharreko sakabanaketan zer-nolako eragina izan zuten ikus daiteke eta migrazio, babes edota desagertze fenomenoak ebalua daitezke.
Taxaceae familiaren agerpena
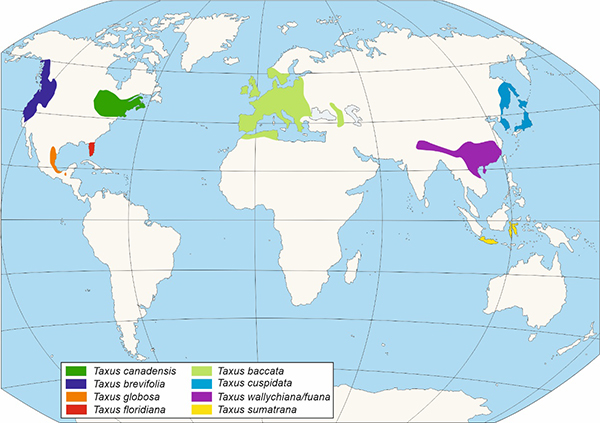
Gimnospermoaren barnean Taxaceae familia gainerako koniferoetatik argi bereizten den multzo bat da. Familia hori, gaur egun, bost generotan irudikatzen da, eboluzioan bi kladotan banatuta. Horietako batean, hiru genero ageri dira, Taxus, Austrotaxus eta Pseudotaxus, eta bigarrengoan bi (Torreya y Amentotaxus), orotara 28 espezie hartzen dituztenak Eurasia, Afrika Iparralde, Kaledonia Berria eta Amerikan (1. Irudia) banatuta. Horiei desagertutako espezie ezberdinak gaineratu behar zaizkie.
Informazio paleoekologikoak klimaren bilakaera eta bioma eta espezie ezberdinengan horrek duen eragina ezagutzeko aukera ematen digu. Horri dagokionez, landare arrastoen ikerketak ekosistemen eta espezieen dinamikaren inguruko ikuspegi zehatzenetako bat ematen digu. Azterketa paleobotanikoek espezie ezberdinek klima aldaketa edo asaldura ezberdinen aurrean duten erantzuna aztertzeko aukera ematen digute.
Zuhaitz espezieetako makrofosilen (enbor, adar, hosto edota fruituen aztarnak) eta mikrofosilen (polen-aleak) azterketatik abiatuz horiek lurralde jakin batean izandako bilakaera berreraiki daiteke. Hala, horietan izandako alterazio natural eta antropikoek denboran zeharreko sakabanaketan zer-nolako eragina izan zuten ikus daiteke eta migrazio, babes edota desagertze fenomenoak ebalua daitezke.
Taxaceae familiaren agerpena
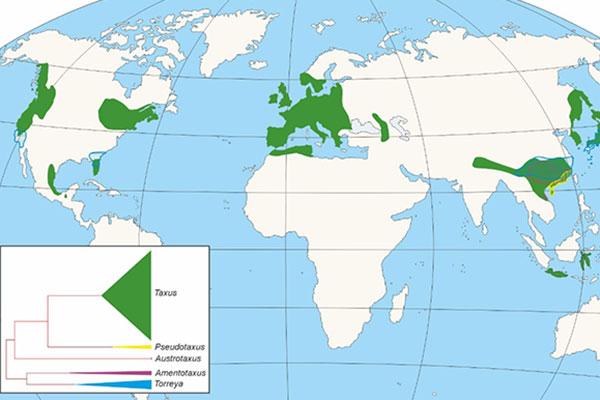
Gimnospermoaren barnean Taxaceae familia gainerako koniferoetatik argi bereizten den multzo bat da. Familia hori, gaur egun, bost generotan irudikatzen da, eboluzioan bi kladotan banatuta. Horietako batean, hiru genero ageri dira, Taxus, Austrotaxus eta Pseudotaxus, eta bigarrengoan bi (Torreya y Amentotaxus), orotara 28 espezie hartzen dituztenak Eurasia, Afrika Iparralde, Kaledonia Berria eta Amerikan (1. Irudia) banatuta. Horiei desagertutako espezie ezberdinak gaineratu behar zaizkie.
1. irudia Taxaceae familiako generoen banaketa eta horietako bakoitzaren agerpena eta garrantzia erakusten duen kladograma.
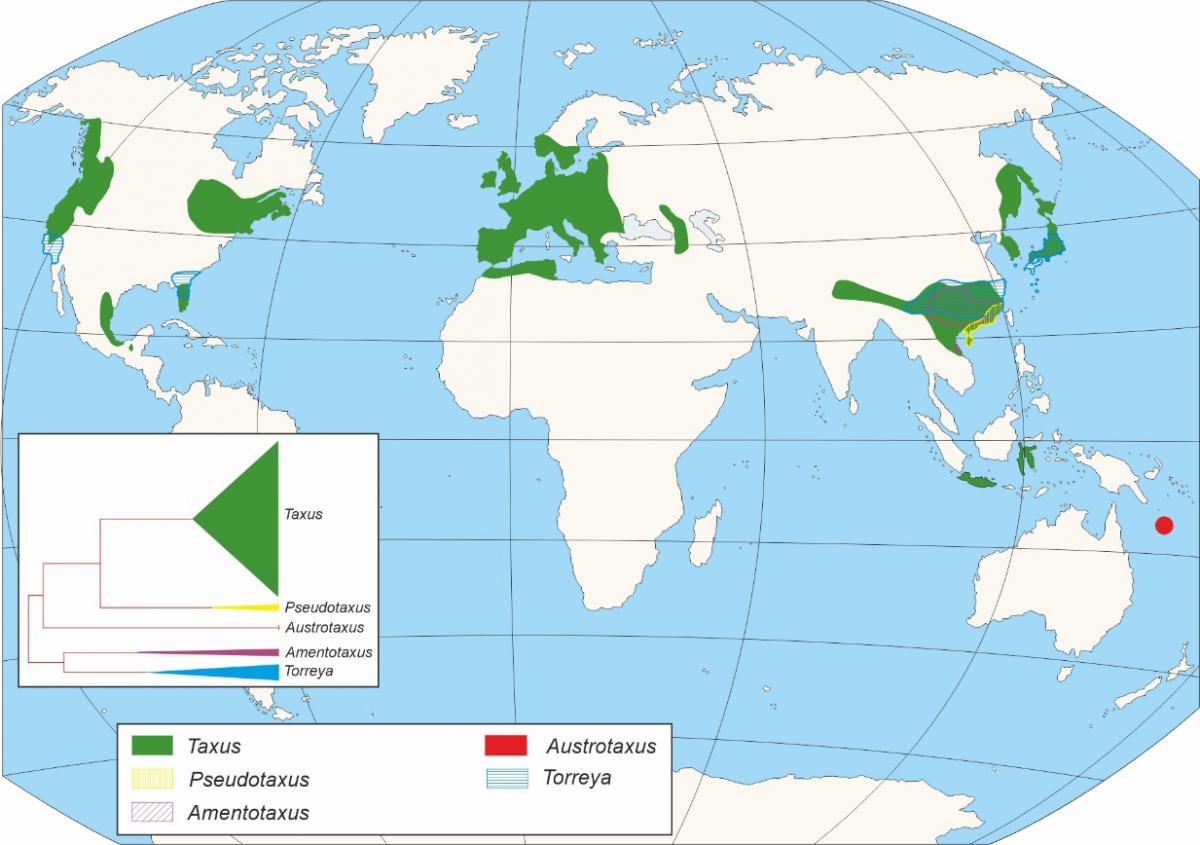
Taxaceae familiaren agerpena Mesozoikoan kokatzen da (2. Irudia), zehazki, Triasiko aldian, duela 250-200 Mu (Mu: milioi urte). Taxaceae Txinako hego-mendebaldetik hedatu zen, Europako leinua osatuko zuten klado ugaritan barreiatuz (1. irudia). Taxaceae familiako antzintasun handieneko fosila Paleotaxus rediviva espezieko hostodun adar baten zati bati dagokio, 200 Mu-ko antzinatasuna duen fazies batetik berreskuratua Suedia hegoaldean (Nathorst 1908). Paleotaxus rediviva horretatik abiatuz Jurasikoan zehar, duela 140 Mu, Taxus jurassica multzoak izango zuen bilakaera (2. irudia). Beranduago, Kretazeo eta Zenozoikoan zehar, espezie berriak ezberdintzen dira, kontinente-unitateen zedarriztatzean zein izandako kliman aldaketa handiek modulatutako prozesu batean. Aldaketa horiek aipatu espezieen sakabanaketan eragina izan zuten eta populazio multzo ezberdinak Ipar hemisferioko lurralde ezberdinetan isolatuta geratu ziren.
2. irudia- Taxus izenekoaren agerpenarekin lotutako jazoera ezberdinen kokapena eskala geologikoan. Gainera, beste jazoera adierazgarri zenbait erakusten dira. Mu: milioi urte.
Taxus generoa (2. eta 3. irudiak) adarkadura irregularreko zuhaitz dioikoez osatzen da, orratz formako hostoak dituztenak eta ezkatadun kimuak, Taxaceae familian garrantzitsuena izanik. Taxus generoa hamar espeziek osatzen dute gutxienez. Dena den, horien kopuruari buruz bada eztabaida bat, nagusiki Asiako kontinentean, eta oraindik ebazteke dagoen kontutzat hartzen da (Hao eta beste, 2008). Taxus generoari atxikitako lehendabiziko aztarna fosilak Taxus jurassica modura izendatu ziren (140 Mu). Horien ezaugarri morfologikoei esker egungo espezie ezberdinekin asimila daitezke: T. baccata, T. cuspidata, T. brevifolia (Florin, 1958).
3. irudia – Taxus generoko espezie ezberdinen banaketa (Li eta beste, 2001).
Aro Mesozoikoa (2. irudia), duela 245 Mu-tik duela 65 Mu-ra bitartean hedatzen da. Aro horretan zehar, Pangea superkontinentea osatzen duen masa kontinentala egonkor mantendu zen, alterazio handirik gabe. Baldintza klimatikoak egonkor mantendu ziren funtsean, eta kontinentean zehar giro tropikal eta subtropikalak nagusitu ziren, banaketa kosmopolitako taxaren nagusitasuna ekarri zuena ondorio lez. Aro horretan zehar nagusitu ziren loredun landareak gimnospermoak izan ziren, horien artean, Taxaceaea eta beste konifero batzuk (esate baterako, ginkgoak eta zikak). Aldiz, Mesozoikoaren amaiera aldera, behe kretazeotik aurrera, angiospermen lehendabiziko fosilak agertzen hasi ziren (2. irudia).
Datu paleobotanikoak kontuan izanik, Taxus da Europako kontinenteko zuhaitz genero zaharrena, ondasun genero jotzen delarik. Taxus fosil zaharrena, Europan berreskuratua, Miozenoaren hastapenean kokatzen da, duela 23 Mu (2. irudia). Taxus engelhardtii izenekoaren hosto batzuekin dator bat, Wiesako udalerrian lokalizatua, Saxonian (Kunzmann & Mai, 2005). Miozenoaren hasierak klimari dagokionez planeta mailan ezegonkortasun aro luze bat dakar. Horrek basoak eta espezie tropikal nagusiak urritzea eta pixkanaka elementu arktotertziarioak eta hostogogorren garrantzia areagotzea eragin zuen. Miozenoaren eta Pliozenoaren arteko trantsizioari dagokionez, makrofosil ezberdinak berreskuratu dira, hazi, aldaxka edo hostoak barne, Europa erdialde eta ekialdeko kokapen sakabanatuetan.
Iberiar Penintsulan, aztarna fosil zaharrenak Banyoles-Besalun, Gironan, Plio-pleistozenoko aintzira-arroaren barnean (Leroy, 2008) eta Galiziako eta Portugal iparralde eta erdialdeko mehatze-arroetan (Ramil-Rego eta beste, 2011) eskuratutako sekuentzia ezberdinetan berreskuratutako polen-aleekin datoz bat. Ingurune bazterreko horietan, Taxus edo Taxaceae zenbait polen-ale isolatu erregistratzen dira, Pliozeneko arloarekin lotzen direnak (duela 5 milioi urte)

4. irudia- Gimnospermoaren enbor fosila (gutxi gorabehera, 150 milioi urte), Montenegro de Andrade Irakasleak jasoa Caldas da Rainhan (Portugal) eta Portoko Unibertsitateko errektoretzako eraikinean erakusgai ipinia.
Pliozenoko klimak euri jasa handiko periodo hotzen, latitude altuenetan oso hotzak izango liratekeenak izotz metaketekin, eta periodo lehor eta beroen arteko txandakatzea du ezaugarri. Periodo horrek Iberiar Penintsulan zuhaitzetako floraren erregistroari dagokionez angiosperma arktotertziarioen nagusitasun erabatekoa, gimnospermoen garapen handi batekin, horien artean kokatuko litzatekeelarik Taxusa, eta elementu tropikalen multzo urri bat, horien artean palmondoen espezie ezberdinak daudelarik, du ezaugarritzat.
Taxusaren dinamika Pleistozenoan zehar
Pleistozenoaren hastapenak, duela 2.4 Mu, aurrerapen glaziarreko ziklo jarraitua, gutxi gorabehera 100.000 urteko iraupena duena, ezaugarritzat duen klima dakar, tartean ontasun handiagoko faseak izanik, 10.000 urteko iraupenarekin. Modu horretan, denbora luze horretan zehar landarediaren garapen zikliko bat ematen da, izaera errepikakorra zuten klima aldaketei erantzunez. Gaur egun, aitortzen da oszilazio klimatiko horien segidak Europako zuhaitz espezieen egungo banaketan eta horien patroi genetikoetan eginkizun erabakigarria bete zutela.
Taxus familiaren dinamikaren ikerketa ziklo glaziarren segida laku edo zohikaztegietako sedimentuetan eskuratutako polenari buruzko zundaketa analisian oinarritzen da funtsean. Europako kontinentean hasierako Pleistozenoko glaziazioartekoetarako eskuragarri dauden datuak sekuentzia zatikatu urri eta sakabanatuetara mugatzen dira. Glaziar-glaziazioarteko azken zikloei dagokienez eskuragarri dagoen informazioa areagotu egiten da oraindik urriak diren eta kontinentean zehar banaketa desberdina agertzen duten sekuentzia jarraituen presentziarekin (6. irudia).
Pleistozenorako eskuragarri dagoen informazio paleobotanikoak agerian uzten duenez, Taxus periodo honen hastapenetik dago Europan. Zentzu horretan, Taxus grandis taxon fosil kuaternarioa, ia milioi bat urteko antzinatasuna duena, Taxus baccata izenekoaren berdin-berdina da ezaugarrietan erreparatuz (Hartzell, 1991). Gainera, Pleistozenoko hasierako glaziazioartekoei esleitutako sedimentu ezberdinetan eskuratutako polenari buruzko datuek Taxusaren polenaren presentzia hasierako Pleistozenotik dagoela erakusten dute, zehazki, Tiglian-C5 glaziazioartekotik, duela 1,8 milioi urte, (Kase, 1988; Deforce & Bastiaens, 2007).
Europa kontinentalerako eta Britainiako uharteetarako eskuragarri dauden polenari buruzko sekuentziek elementu horren garrantzia Pleistozeno ertaineko glaziazioarteko jarraituak igarotzen diren heinean areagotuz doala erakusten dute, paisaia aldetiko garrantzia handiena Holsteinian glaziazioartekoan hartuz (Jessen eta beste, 1959, West, 1962, Kelly, 1964, Godwin, 1975, Watts, 1985, Coxon, 2017), duela 400–367 ka (ka: milaka urte). Glaziazioarteko horrekin lotuta 1911n Clacton-on-Sea herrian (Essex, Erresuma Batuak), berreskuratutako Taxuz zurezko lantza (Godwin 1975, Oakley eta beste, 1977) kokatzen da, Europan zurarekin fabrikatutako artefaktu zaharrenetakoa dena.
Iberiar Penintsulan, Taxusari dagokionez, Pleistozenoko polenaren aztarna antzinakoena Bobila-Ordis lakuko erregistro sedimentologikoan eskuratutako erregistroari dagokio (Leroy, 2008), Ipar-ekialdean lekututa. Erregistroa Pleistozenoaren hastapeneko glaziazioarteko periodo batekin lotzen da, nahiz eta ikerketan ez den zehazten hasierako Pleistozenoko zein periodo interrestadiali dagokion. Iberiar Penintsulan Taxusaren presentziari buruzko beste ebidentzia batzuk Penintsulako erdialdeko zonan berreskuratutako hiena koprolitoetan identifikatutako polen-aleekin bat datoz, Pleistozeno Ertainarekin lotzen direnak (Carrión eta beste, 2006).
Azken ziklo Glaziar - Glaziazioartekoa Iberiar Penintsularen Iparraldean
Duela, gutxi gorabehera, 110.000 urte, abiatu zen azken ziklo glaziar - glaziazioartekoa (5. Irudia), Eemiense glaziazioartekoarekin (duela 115-95 milaka urte) eta Würm glaziarrarekin (duela 95-15 milaka urte). Europan periodo horretarako eskuragarri dagoen informazio paleobotanikoa areagotu egiten da, lurraldearen araberako irudikapen hobea lortuz, kontinentean barna sakabanatua izan arren (6. irudia).
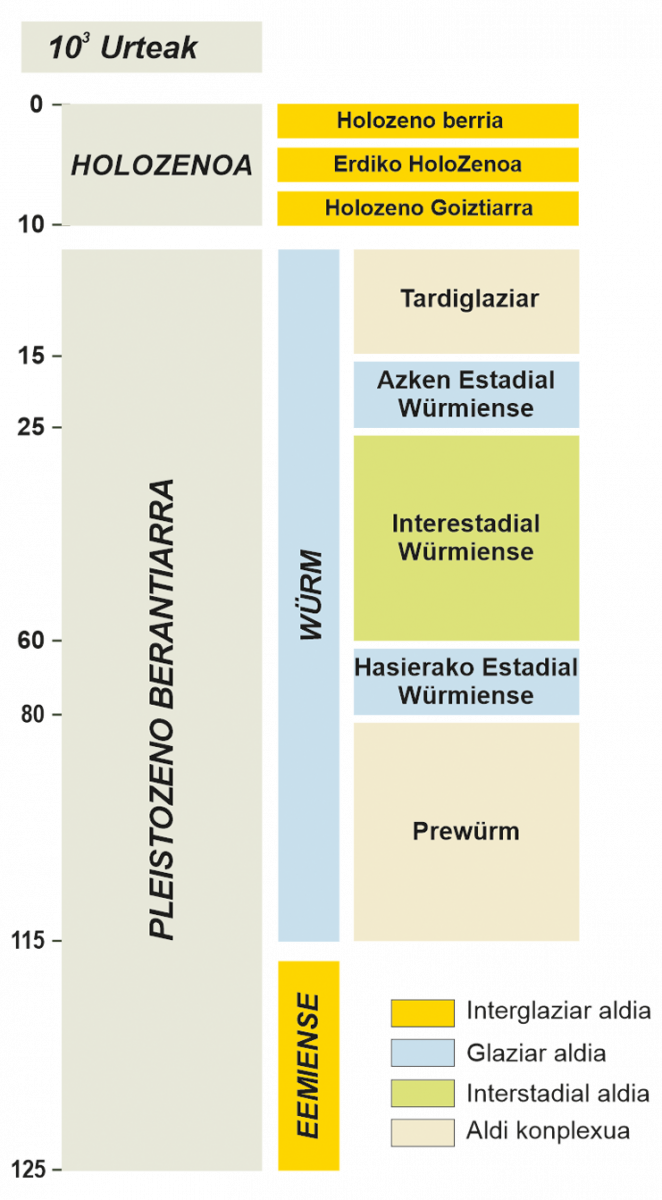
5. irudia.- Azken ziklo glaziar - glaziazioartekoaren periodizazioa Europako hego-mendebaldean
Azken glaziazioartekoan zehar (Eemiense) Europako kontinenterako periodo horri dagokionez eskuragarri dauden sekuentzia gehienek Taxusaren hedapenaren fase adierazgarri bat erakusten dute, nagusiki Europa erdialdeko lurretan, eta haren garrantzia murriztuz ekialdean eta hegoaldean, penintsula balkaniko eta italikoetako zenbait eremutan agertu ere ez delarik egiten. Iberiar Penintsulan glaziazioarteko horri dagokion informazio paleobotanikoa oso eskasa da. Taxusaren presentzia erakusten duen datu bakarra isolatutako polen-ale batzuen erregistroa da Paduleko zohikaztegiko sekuentzia Eemiensean, Granadan (Camuera eta beste, 2019).
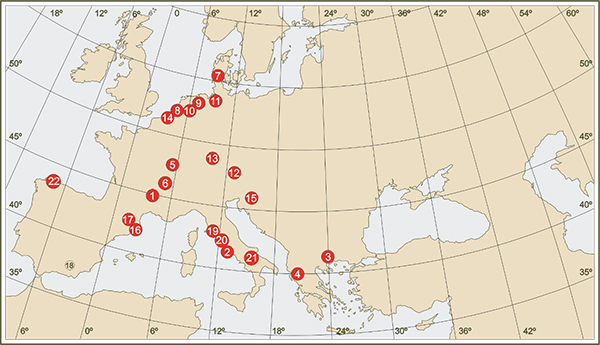
6. irudia.- Pleistozenoko ziklo glaziar bat baino gehiago (1- Bouchet/Placraux, 2- Valle di Castiglione, 3-Tenaghi Phillipon, 4- Ioannina) edo azken ziklo glaziar osoa edo zati handi bat (5- La Grande Pile, 6- Les Echets, 7- Brorup, 8- Amersfoort, 9- Denekamp, 10- Hengelo, 11- Oerel, 13- Samerberg, 14- Moershoofd, 15- Ljubljana Moor, 16- Banyoles. 17- Pla de l’Estany, 18- Padul, 19- Lagaccione, 20- Lago di Vico, 21- Lago Grande di Monticchio, 22- Arealonga) erregistratzen duten Europako kontinenteko polenari buruzko sekuentzia nagusien kokapena.
95.000-15.000 urte bitarteko periodoan (5. irudia) Ipar hemisferioan klima asko hoztu zen. Horren ondorioz, paisaian eta baso flora eta faunaren etorkien banaketan eta bilakaeran aldaketa handiak izan ziren. Mendialdeko lurretan, oro har 900-1.100 metrotik gora, baldintzak zinez hotzak ziren eta izotzak ziren nagusi, zuhaitz landaredia mantentzeko baldintza bateraezinak. Izotzak modu iraunkorrean edo aldi baterako estalitako eremuen azpitik paisaia zuhaitzik gabekoa zen erabat. Eremu atlantikoetan belar-formazioak ezartzen dira, non gramineak nagusitzen diren. Eremu kontinentaletan, ordea, hezetasun gabeziak etapa lehorren nagusitasuna baldintzatzen du. Belar-formazioen azpitik edo horiekin elkarlotuak lurraldearen berezitasunetara egokitutako mosaiko ezberdinetan, zuhaixka erako formazioak, erikoideen formazioak eta leguminoso arantzatsu eta arantzagabeen formazioak lekutzen dira eremu Atlantikoetan (Txilardiak). Aitzitik, eremu kontinentaletan Juniperus (Juniperus communis, Juniperus sabina) erako formazioak dira nagusi, leguminoso pulbilunarrekin batera. Paisaian nagusitzen diren belar- eta zuhaixka-formazioek ornodun handi, ertain eta txikien fauna adierazgarria batzen dute. Basoak lurraldearen eremu babestuenetan itxita geratzen dira, eta berriro banaketa bat aurki dezakegu eremu Atlantikoen eta eremu kontinentalen artean. Hala, lehendabizikoetan angiosperma hostoerorkorrak (Quercus, Alnus, Betula, Corylus, Castanea, Fagus, etab.) dira nagusi eta hostoiraunkorrak ere badira. Bigarrenetan, aldiz, Pinus direlakoak gailentzen dira, eta angiospermen presentzia urriagoa da.
Iberiar Penintsularen ipar-mendebaldean, azken periodo glaziarrerako informazio paleobotanikoa oso urria da, egungo kostaldeko lerroan kokatutako sekuentzia segida batera mugatuz. Eskuragarri dauden datuetan ez da Taxusaren presentziarik erregistratzen. Würm glaziarrean zeharreko Taxusaren polenaren erregistrorik eza errepikatu egiten da hala Iberiar Penintsulako beste eremu batzuetan nola Europako kontinentean barna.
Taxusaren dinamika Tardiglaziarrean zehar
Azken ziklo Glaziar-Glaziazioartekoaren amaierako etapetan tenperatura igo egin zen orokorki. Horrek eragina izan zuen paisaiaren konfigurazioan eta egungo glaziazioartekoa ekarri zuen, Holozenoa, duela 10.000 urtetik gaurdainora hedatzen dena (7. irudia). Tardiglaziar izeneko periodo horretan zehar, lur eta itsasoko azalera zabalak estaltzen zituzten izotzen fusioa gertatu zen. Izotzen gailentasuna murriztu egin zen, Holozenoaren erdialdera altuera handieneko tontorretara mugatzen zelarik. Baldintza klimatikoak hobetu egin ziren eta, horren ondorioz, basoak hedatu. Zuhaitzen formazioak gero eta handiagoak ziren eta gero eta lur gehiago hartuz zihoazen, Holozenoaren erdialdera hedadura handiena izan arte, klima holozeno egokiarekin bat etorriz. Iberiar Penintsularen ipar-mendebaldean angiospermen formazio hostoerorkorrak nagusitu egin ziren kantauriar-atlantikoko isurialdeko eremu ozeanikoenetako zati handi batean. Aldiz, barneko lurretan pinudiak edo pinudien mosaikoak, angiosperma hostoiraunkorren basoak eta baso hostoerorkorrak nagusitu ziren.
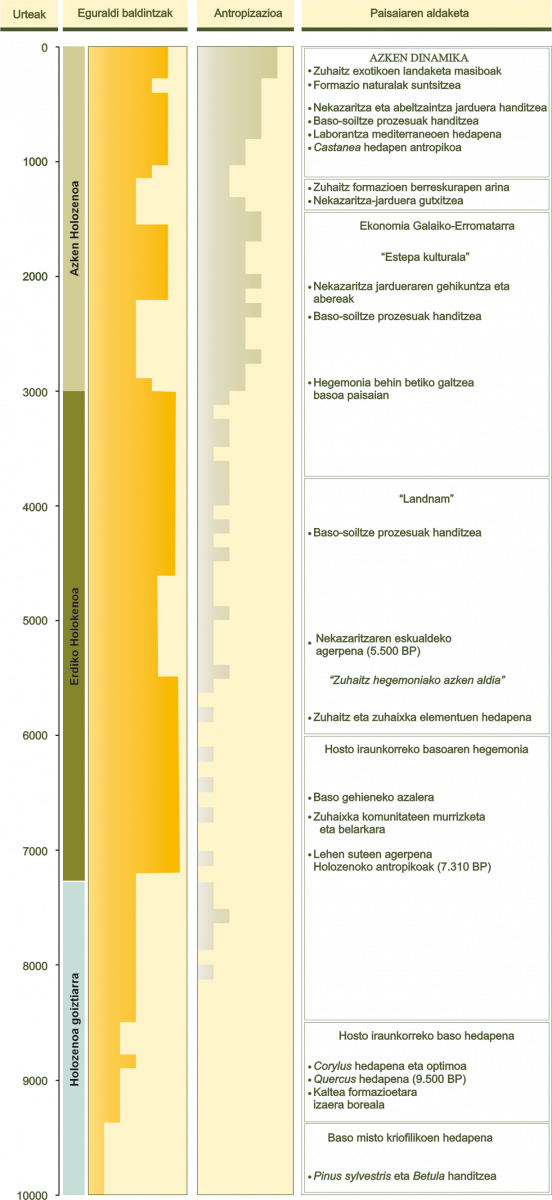
7. irudia.- Holozenoaren ingurumen periodizazioa Galizian.
Deglaziazio tardiglaziarreko faseetan zehar eta egungo glaziazioartekoaren hasieran Iberiar Penintsulako ipar-mendebaldeko diagrametan Taxusaren presentzia urria da. Aldiz, espeziearen iraunkortasunaren lekukotzak aurkitzen dira lurraldean azken ziklo Glaziar-Glaziazioartekoan zehar, egun hartzen dituen kokapenekin alderatuz oso leku ezberdinak hartuz, izan ere, gaurko horiek, gehienean, Pleistozenoan zehar espeziearen garapenarekin bateraezinak ziren baldintzak agertzen zituzten. Desizozte periodo horretan zehar Taxusaren presentzia erregistratzen da Pena Vellan, Galiziako iparraldeko mendilerroetan (Ramil-Rego, 1992), La Royan (Allen eta beste, 1986), Sanabrian eta Quintanar de la Sierran (Peñalba, 1989) Burgosen (1. taula, 8. irudia).
Taxusaren dinamika Holozenoaren hasierako faseetan
Haginaren lurraldekako banaketa areagotu egin zen Holozenoan zehar, altueran gero eta gehiago agertuz Iberiar Penintsularen iparraldeko eskualde ezberdinetan (1. taula, 6. irudia). Holozenoaren ezaugarri diren basoaren hedapen eta gailentasunaren faseetan zehar, Taxusen erregistro kopurua areagotu egin zen Penintsula Iberikoko ipar-mendebaldean. Modu horretan, kantabriar isurialdean zehar presentzia ezberdinak erregistratzen dira, hala polen-aleenak polen-sekuentzietan, nola ikatzenak aztarnategi arkeologikoetan (1. eta 2. taulak) Bi kasuetan, eskuragarri dauden datuek haginen presentziarekin bat etorriko liratekeen unean uneko presentziak aipatzen dituzte, nagusi ziren zuhaitz masen barnean sartutako indibiduo isolatu modura, hagin multzoen presentziaren zantzurik hauteman gabe.
| Gordailua | Tr | HH | HE | HB | Erreferentzia |
| Pena Vella | Ramil Rego, 1992 | ||||
| Pozo do Carballal | Muñoz Sobrino, 2001 | ||||
| La Roya | Allen et al., 1996 | ||||
| Cueto de Avellanosa | Núñez de la Fuente, 2018 | ||||
| Zalama | Pérez Díaz et al., 2016 | ||||
| Los Tornos | Peñalba, 1989 | ||||
| Los Tornos 3-5 | Peñalba, 1989 | ||||
| Quintanar de la Sierra | Peñalba, 1989 | ||||
| Belate | Peñalba, 1989 | ||||
| Atxurri | Peñalba, 1989 | ||||
| Saldropo 1 | Peñalba, 1989 | ||||
| Inurritza 1 | Peñalba, 1989 | ||||
| Fuente del Vaquero | Pérez-Díaz, 2012 |
1. taula.- Iberikoko ipar-mendebaldeko polen-sekuentziak, non Taxusaren polenaren presentzia identifikatu den (Tr:: Tardiglaziarra, HH: Hasierako Holozenoa, HE: Holozeno Ertaina, HB: Holozeno Berria).
| Gordailua | HH | HM | HB | Erreferentzia |
| Atxoste | Ruíz Alonso, 2003-2007 | |||
| Buruntza | Olaetxea, 1997 | |||
| Ondarre | Mujika et al., 2013 | |||
| Mulisko Gaina | Peñalba, 1987 | |||
| Aizpea | Zapata, 2001 | |||
| Peña Larga | Zapata, 2001 | |||
| Arrubi | Ruiz-Alonso, 2003-2007 | |||
| Mendandia | Iriarte, 2006 | |||
| Peña Parda | Ruiz-Alonso & Zapata, 2003 | |||
| El Mirón | Zapata, 2012 | |||
| Mazaculos | Uzquiano, 1995 | |||
| Aramo II | Beato et al., 2019 | |||
| Peña Oviedo | Carrión Marco, 2005 | |||
| Pala da Vella | Carrión Marco, 2003 |
2. Taula.- Iberikoko ipar mendebaldeko gordailuak, non Taxusaren ikatzen presentzia identifikatu den(HH: Hasierako Holozenoa, HE: Holozeno Ertaina, HB: Holozeno Berria).
Holozenoaren hasierako faseetan gizakien perturbazioak urriak dira eta eremu geografiko jakin batzuetan metatzen ziren, azken Holozenoan areagotu eta hedatu baziren ere, bereziki Europa mendebaldeko lurretan, non ekosistema naturalak nabarmen murriztu eta isolatu ziren antropoekosistemen aurrean.
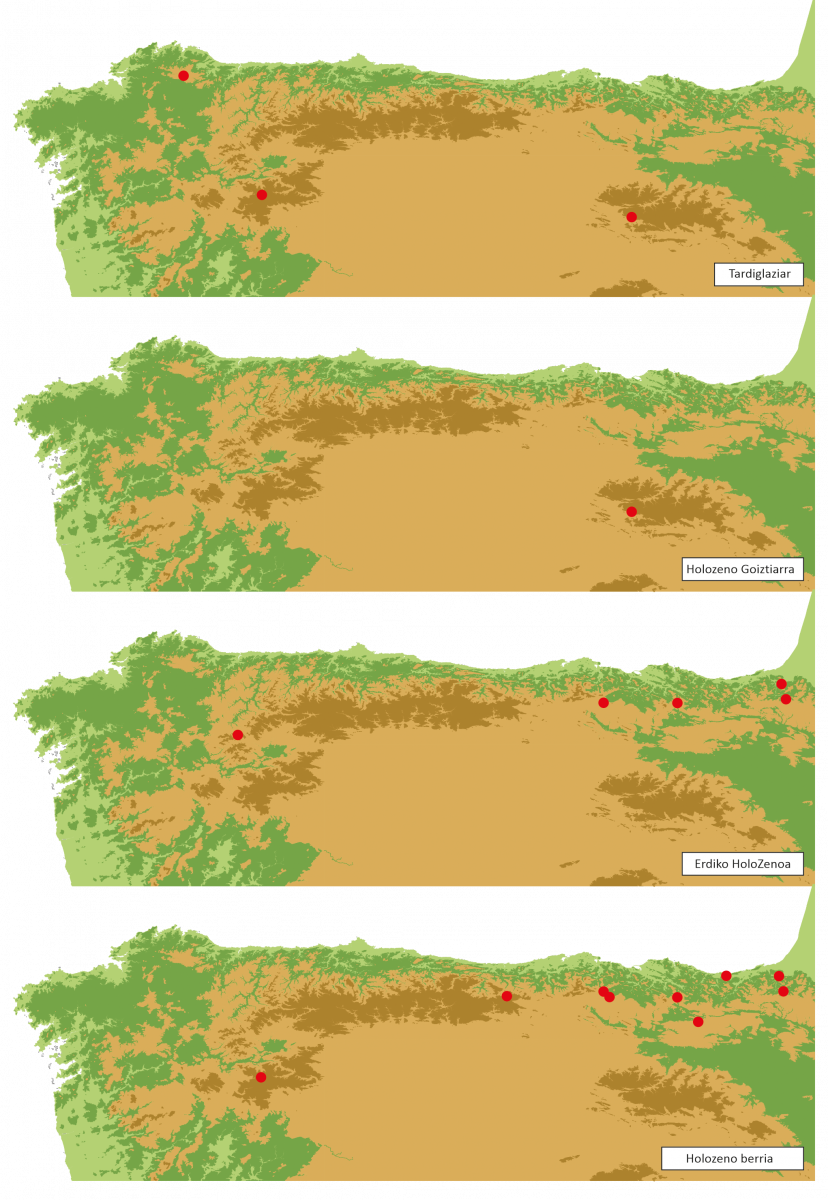
8. irudia.- Taxus polenaren presentzia ageri duen polen-sekuentzian Tardiglaziar eta Holozenoan zehar Iberiar Penintsulako ipar-mendebaldean.
Taxusaren dinamika Holozenoaaren azken herenean
Holozenoaren azken herenean polen-sekuentzietan Taxusaren polenaren agerpena erregistratzen da sekuentzia batzuetan, beste batzuetan, aldiz, haien erregistroa galdu egiten da (8. irudia). Taxusaren polen kopuru handiagoaren erregistroa, Holozenoaren aurreko etapekin alderatuz, nagusi ziren zuhaitz-formazioetako polen-euriaren murrizketarekin lotuko litzateke, sekuentzia askotan baso ugarienen artean modu isolatuan agertzen diren zuhaitz-elementuetako polena ezkutatuko lukeena. Basoak paisaian zuen garrantzia murriztuta, gutxiengoa ziren elementu horiek errazago hautemango lirateke sekuentzietan.
Nekazaritza eta abeltzaintzako ekoizpen-sistemak agertzean funtsezko aldaketak etorri ziren paisaiaren konfigurazioari dagokionez Europa osoan. Iberiar Penintsularen iparraldean prozesu hori duela 5.500 urte hasi zen, landatutako landareen lehendabiziko erregistroak erregistratzen direnean. Brontze Aroaren amaierarekin eta Burdin Aroaren hasierarekin bat etorriz, nekazari eta abeltzainen herrixka txikietan ekoizpena areagotu egin zen, eta baita paisaiaren gainean haiek utzitako arrastoa ere. Modu horretan, duela 2.000 urte gehien populatutako eremuetan, basoak eta landaredia naturala alde batera geratzen hasi zen pertsonen eta etxeko abereen indarrak sostengatutako nekazaritza sistemen konfigurazioaren ondorioz.
Hagina urritzen hasi zen paisaian, eta beste zuhaitz espezie batzuekin gertatu bezala, mendialdean gordean geratu zen, nekazaritza eta abeltzaintzaren presioa horren handia ez den lekuetan. Nekazaritza sistemek konfiguratutako paisaia irekiak mantendu egin ziren eta indartu Erdi Aroan eta Aro Modernoan, baso-eremuen isolamendua mendialdeko zonetara areagotuz. Gune askotan, bereziki mendian, haginak protagonismo berezia hartu zuen garaiotan baliabide medikuntzako, sinboliko eta espiritual modura erabiltzearekin lotuta, etxeko eta erlijio espazioetara sartuz. Bertan iragan konplexu eta urruneko lekuko bizi modura mantentzen dira, formazio natiboak lausotu eta azkenik desagertu egiten diren arte.
Haginaren formazio eta basoak oinarrizko osagaia dira Europako lurraldeko biodibertsitatean, milaka urteetan zehar errotua aldaketa biogeografiko eta klimatiko handien eraginez. Baso eta formazio horiek eremu askotan, Iberiar Penintsulako Eskualde Atlantikoan gertatzen den eran, tokiko komunitateekin harreman hertsia mantendu dute eta mantentzen dute oraindik, azken horiek lehendabizikoek eskaintzen dituzten ekosistema-zerbitzuen onura jasotzen dutelarik.
Bibliografia
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Beato Bergua, S., Poblete Piedrabuena, M.A., Cunill Artigas, R. (2019). Taxus baccata en la Sierra del Aramo (Macizo Central Asturiano). Boletín de la Asociación de Geógrafos Españoles, 81, 2772, 1–30.
Camuera, J., Jiménez-Moreno, G., Ramos-Román, M.J., García-Alix, A., Toney, J.L., Anderson, R.S., Jiménez-Espejo, F., Bright, J., Webster, C., Yanes, Y., Carrión, J.S. (2019). Vegetation and climate changes during the last two glacial-interglacial cycles in the western Mediterranean: A new long pollen record from Padul (southern Iberian Peninsula),Quaternary Science Reviews, 205: 86-105.
Carrión Marco, Y. (2003). El paisaje vegetal prehistórico de Pala da Vella: primeros resultados antracológicos. In: V. Oliveira Jorge. Contributos das ciencias e das tecnologias para a arqueologia da Península Ibérica: 21-32. UTAD Vila Real, Portugal
Carrión Marco, Y. (2005). El impacto de la economía productora en el paisaje vegetal de Peña Oviedo (Camaleño, Cantabria). In: R. Ontañón Peredo, C. García-Moncó Piñeiro, P. Arias Cabal. Actas del III Congreso del Neolítico en la Península Ibérica: 35-44. Santander.
Carrión, J. S., Scott, L., Arribas, A., Fuentes, N., Gil‐Romera, G. and Montoya, E. (2006). Pleistocene landscapes in central Iberia inferred from pollen analysis of hyena coprolites. Journal of Quaternary Science, 22: 191–202.
Coxon, P., McCarron, S., Mitchell, F. (Eds) (2017). Advances in Irish Quaternary Studies. Atlantis Advances in Quaternary Science. Atlantis Press, Paris. 316pp.
Deforce, K. & Bastiaens, J. (2007). The Holocene history of Taxus baccata (yew) in Belgium and neighbouring regions. Belgian Journal of Botany, 140(2): 222-237.
Florin, R. (1958). On Jurassic taxads and conifers from North-Western Europe and Eastern Greenland. Acta Horti Bergiani Series, 17: 257-402.
Godwin H. (1975). The history of the British Flora. A factual basis for phytogeography. Cambridge University Press, Cambridge. 383 pp.
Hao, D.-C., Huang, B., Yang, L. (2008). Phylogenetic Relationships of the Genus Taxus Inferred from Chloroplast Intergenic Spacer and Nuclear Coding DNA. Biological & pharmaceutical bulletin, 31(2): 260-265.
Hartzell, J.R. (1991). The Yew Tree: A Thousand Whispers. Hulogosi Communications, Oregon, 319 pp.
Heim J. (1970). Les relations entre les spectres polliniques récents et la végétation actuelle en Europe occidentale. Ph.D. thesis, Université catholique de Louvain, Louvain-la-Neuve, Belgium.
Iriarte, M.J. (2006). El entorno vegetal del abrigo de Mendandia y su depósito arqueológico: Análisis palinológico. In: Alday A. (Dir.). El legado arqueologico de Mendandia. Los modos de vida de los últimos cazadores en la Prehistoria de Treviño: 405-418. Junta de Castilla y León. Burgos.
Jessen, K., Andersen, S.Th., Farrington, A. (1959). The interglacial deposit near Gort, Co. Galway, Ireland. Proceedings of the Royal Irish Academy B, 60: 1-77.
Kasse K. (1988). Early-Pleistocene tidal and fluviatile environments in the southern Netherlands and northern Belgium. Ph.D. thesis, Vrije Universiteit, Amsterdam, The Netherlands.
Kelly, M.R. (1964) The Middle Pleistocene of North Birmingham. Philos. T. Roy. Soc. B, 247: 533-592.
Kunzmann L., Mai D.H. (2005) Die Koniferen der Mastixioideen-Flora von Wiesa bei Kamenz (Sachsen, Miozän) unter besonderer Berücksichtigung der Nadelblätter. Palaeontographica B 272(1–6): 67–135.
Leroy, S.A.G. (2008): Vegetation cycles in a disturbed sequence around the Cobb-Mountain subchron in Catalonia (Spain). Journal of Paleolimnology, 40(3), 851-868
Li, J., Davis, C.C., Del Tredici, P., Donoghue, M.J. (2001). Phylogeny and biogeography of Taxus (Taxaceae) inferred from sequences of the internal transcribed spacer region of nuclear ribosomal DNA. Harv. Pap. Bot. ,6:267–274.
Mujika-Alustiza, J.A., Agirre-García, J., Edeso-Fito, J.M., Lopetegi-Galarraga, A., Pérez-Díaz, S., Ruiz-Alonso, M., Tarriño-Vinagre, A., Yusta-Arnal, I. (2013). La continuidad de la actividad pastoril durante la época romana en la zona de Argarbi (sierra de Aralar - Gipuzkoa). Kobie Serie PaleoantroPología, 32: 217-258
Muñoz Sobrino, C. (2001). Cambio climático y dinámica del paisaje en las montañas del noroeste de la Península Ibérica. Tesis Doctoral. Escuela Politécnica Superior. Universidad de Santiago de Compostela.
Nathorst A.G. (1908). Palaeobotanische Mitteillungen.7. Über Palyssya, Stachyotaxus und Paleotaxus. Ksvenka Vetensk Akad Handl 8: 1–16.
Nuñez de la Fuente, S. (2018). Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje atlántico montañoso: la región cantábrica durante el Holoceno. Tesis doctoral. Universidad de Cantabria.
Peñalba Garmendia, M.C. (1989). Dynamique de Végétation Tardiglaciaire et Holocene Du Centre-Nord de l’Espagne d’après l’analyse pollinique. These. Université d’Aix, Marseille III.
Pérez-Díaz, S. (2012). El paisaje vegetal durante la Prehistoria Reciente en la vertiente mediterránea de Euskal Herria— Tesis doctoral. Euskal Herriko Univ., Vitoria-Gastei.
Perez-Díaz, S., López-Sáez, J. A., Pontevedra-Pombal, X., Souto-Souto, M. & Galop, D. (2016). 8000 years of vegetation history in the northern Iberian Peninsula inferred from the palaeoenvironmental study of the Zalama ombrotrophic bog (Basque-Cantabrian Mountains, Spain). Boreas, Vol. 45, pp. 658–672.
Ramil Rego P, Gómez-Orellana L, Muñoz Sobrino C, Tereso JP (2011). Dinâmica natural e transformação antrópica das florestas do noroeste ibérico. In Tereso JP, Honrado JP, Pinto AT, Rego FC (Eds.) Florestas do Norte de Portugal: História, Ecologia e Desafios de Gestão. InBio - Rede de Investigação em Biodiversidade e Biologia Evolutiva. Porto. ISBN: 978-989-97418-1-2. Pp 14-54.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ruiz-Alonso, M. & Zapata, L. (2003). Análisis antracológico del yacimiento arqueológico de Peña Parda. Cuad. Arq. Univ. Navarra, 11: 217-252.
Ruiz-Alonso, M. (2003/2007). Madera carbonizada en los fondos de cabaña de Arrubi y Esnaurreta (Sierra de Aralar, Gipuzkoa): vegetación y recursos forestales en la Edad Media. Kobie (Serie Paleoantr.), 27: 131-150.
Uzquiano, P. (1995). L’evolution de la vegetation a l’Holocene inicial dans le nord de l’Espagne a partir de l’étude anthracologique de trois sites archeologique. Quaternaire 6 (2): 77-83.
Van der Knaap, W.O., van Leeuwen, J.F.N. (1997). Late Glacial and early Holocene vegetation succession, altitudinal vegetation zonation, and climatic change in the Serra da Estrela, Portugal. Review of Palaeobotany and Palynology, 97:3-4: 239-285.
Watts, W.A. (1985) Quaternary vegetation cycles.In: Edwards, K.J. & Warren, W.P. (eds.): The Quaternary history of Ireland: 154-185. Academic Press, London.
West, R.G. (1962) A note on Taxus pollen in the Hoxnian Interglacial. New Phytologist, 61: 189-190.
Zapata, L. (2012). Holocene wood charcoal from El Mirón cave. Vegetation and wood use — In: Straus, L.G. & González Morales, M.R. (Eds.). El Miron cave. Cantabrian Spain. The site and its Holocene archaeological record: 174-196. Univ. New Mexico, Alburquerque.