


History of yew forests in Northern Iberian Peninsula
16-12-2020
The earth over time has undergone numerous transformations that conditioned life on the planet. The main changes in the earth history were continental drift and climate change. The most important force of climate change in the past was the modification of the earth's orbital cycles, which ultimately determine the amount of insolation that each part of the planet receives and therefore the climate in the different areas of the globe. However, the last years of the Earth's history are marked by modifications derived from human alteration in natural ecosystems.
Paleoecological information allows us to know the evolution of the climate and its influence on the different biomes and species. In this sense, the study of plant remains provides one of the most accurate views of the ecosystem and species dynamics. Paleobotanical analyses allow us to analyse the response of different species to climatic changes or other disturbances.
From the study of the macro-fossils (remains of trunks, branches, leaves, fruits, etc.) and micro-fossils (pollen grains) of the tree species, their evolution in a given territory can be reconstructed, observing how the different natural and anthropogenic changes influenced their distribution over time and evaluating the phenomena of migration, refuge, extinction, etc.
Appearance of Taxaceae
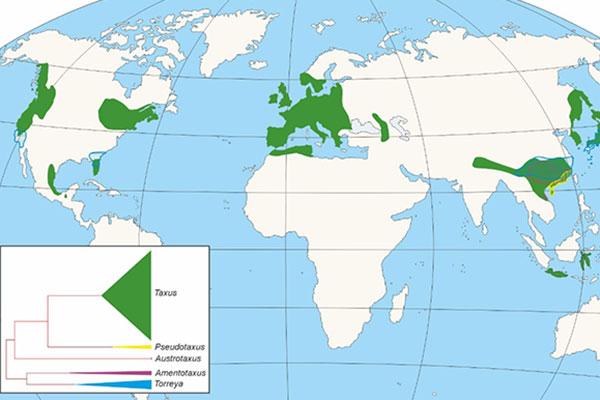
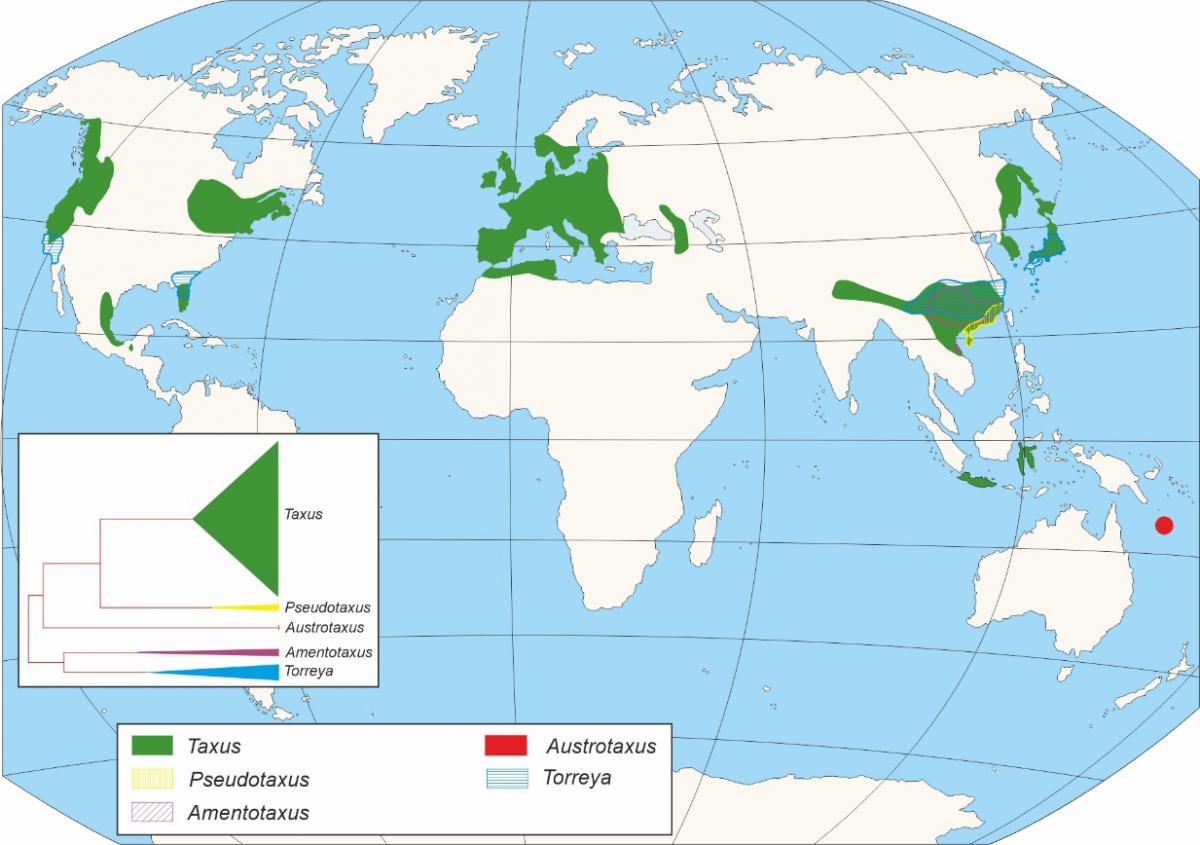
Within the gymnosperms, Taxaceae family constitutes a group clearly differentiated from the rest of conifers, being currently represented by five genera evolutionarily divided into two clades, one with three genera Taxus, Austrotaxus and Pseudotaxus, and the second with two (Torreya and Amentotaxus), which together collect about 28 species distributed in Eurasia, North Africa, New Caledonia and America (Figure 1), to which different extinct species must be joined.
Paleoecological information allows us to know the evolution of the climate and its influence on the different biomes and species. In this sense, the study of plant remains provides one of the most accurate views of the ecosystem and species dynamics. Paleobotanical analyses allow us to analyse the response of different species to climatic changes or other disturbances.
From the study of the macro-fossils (remains of trunks, branches, leaves, fruits, etc.) and micro-fossils (pollen grains) of the tree species, their evolution in a given territory can be reconstructed, observing how the different natural and anthropogenic changes influenced their distribution over time and evaluating the phenomena of migration, refuge, extinction, etc.
Appearance of Taxaceae
Within the gymnosperms, Taxaceae family constitutes a group clearly differentiated from the rest of conifers, being currently represented by five genera evolutionarily divided into two clades, one with three genera Taxus, Austrotaxus and Pseudotaxus, and the second with two (Torreya and Amentotaxus), which together collect about 28 species distributed in Eurasia, North Africa, New Caledonia and America (Figure 1), to which different extinct species must be joined.
Figure 1. Distribution of Taxaceae genera and cladogram showing the appearance and importance of each one of them.
The appearance of Taxaceae family comes from the Mesozoic (Figure 2), specifically in the Triassic period, about 250-200 My (My: million years). Taxaceae radiated from southwest China, diverging into several clades that would include the European lineage (Figure 1). The oldest fossil of Taxaceae corresponds to a fragment of a branch with leaves of Paleotaxus rediviva recovered in a 200 My facies in 1908 in southern Sweden (Nathorst 1908). From Paleotaxus rediviva, the group Taxus jurassica would evolve during the Jurassic, 140 My ago (Figure 2). Later, during the Cretaceous and Cenozoic, new species were differentiated, in a process that was modulated by important changes both in the delimitation of the continental units and in the climate, which affected their distribution and the isolation of different population groups in different territories of the Northern hemisphere.
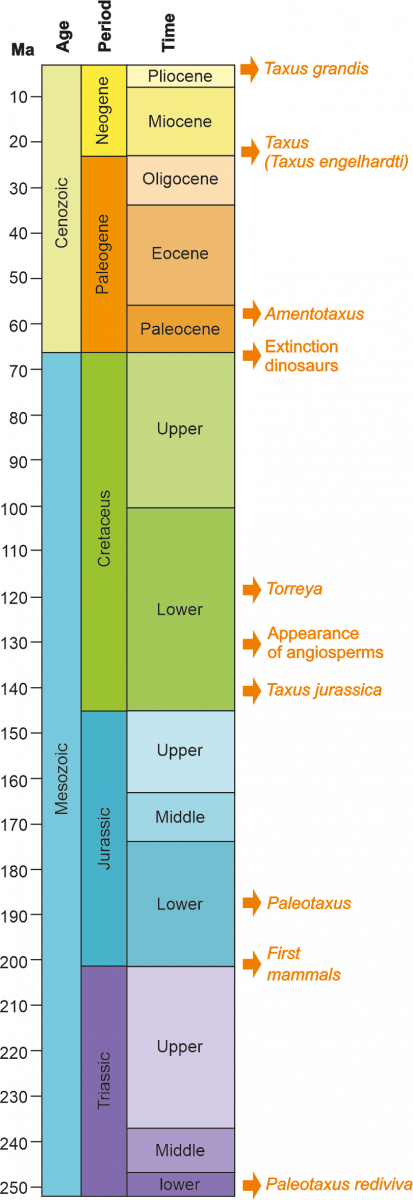
Figure 2.- Location on a geological scale of the different events related to the appearance of Taxus. In addition, other significant events are displayed. My: millions of years.
The Taxus genus (Figures 2 and 3) is made up of dioecious trees with irregular branching, with acicular leaves and scaled buds, being the most important of the Taxaceae family. The Taxus genus is made up of at least ten species, although there is a debate regarding the number of them, mainly in the Asian continent, considering it a still unresolved issue (Hao et al., 2008). The first fossil remains assigned to the Taxus genus were designated as Taxus jurassica (140 Ma). The morphological characteristics of these remains allow their assimilation with different current species: T. baccata, T. cuspidata, T. brevifolia (Florin, 1958).
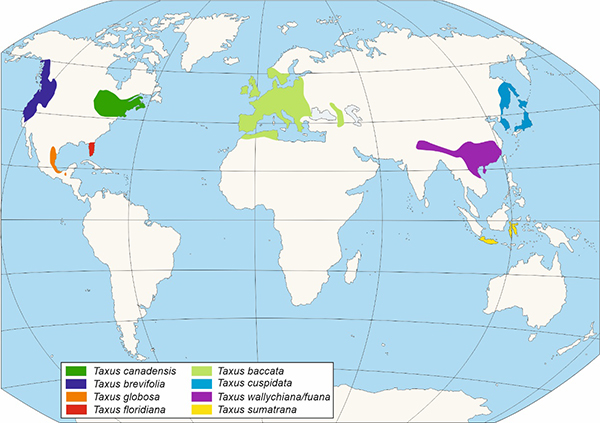
Figure 3. - Distribution of the different species of the Taxus genus according to Li et al., 2001
.
.
The Mesozoic era (Figure 2) stretches between from 245 My to 65 My. During it, the continental mass that constituted the Pangea supercontinent remains stable without undergoing major alterations. The climatic conditions remained basically stable, prevailing the tropical and subtropical environments that would mark the predominance of the taxa of cosmopolitan distribution throughout the continent. The dominant flowering plants during this era are gymnosperms, which would include Taxaceae and other conifers (such as ginkgos and cycas). In the final part of the Mesozoic, from the Lower Cretaceous, the first fossils of angiosperms begin to appear (Figure 2).
Taking paleobotanical data into account, Taxus is the oldest genus of trees on the European continent, being considered a relict genus. The oldest Taxus fossil, recovered in Europe, dates from the early Miocene, about 23 Ma (Figure 2). It corresponds to some leaves of Taxus engelhardtii located in the town of Wiesa in Saxony (Kunzmann & Mai, 2005). The beginning of the Miocene determines the beginning of a long stage of climatic instability at a planetary level, which will mark the reduction of the dominant tropical forests and species and the gradual increase in importance of the arctoterciary and sclerophilic elements. In relation to the transition between Miocene and Pliocene, different macro-fossils, including seeds, twigs or leaves, have been recovered in scattered locations in Central and Eastern Europe.
In the Iberian Peninsula, the oldest fossil remains correspond to pollen grains recovered in various sequences obtained within the Plio-Pleistocene lake basin of Banyoles-Besalú, in Girona (Leroy, 2008) and in the mining basins of Galicia, North and Central Portugal (Ramil-Rego et al., 2011). In these depositional environments, some isolated pollen grains of Taxus or Taxaceae are recorded, related to the Pliocene era (5 million years ago)
 Figure 4.- Gymnosperm fossil trunk (approx. 150 million years old), collected by Professor Montenegro de Andrade in Caldas da Rainha (Portugal) and exposed in the building of the Universidade do Porto’s Rectorate.
Figure 4.- Gymnosperm fossil trunk (approx. 150 million years old), collected by Professor Montenegro de Andrade in Caldas da Rainha (Portugal) and exposed in the building of the Universidade do Porto’s Rectorate.The Pliocene climate is characterized by the alternation of cold periods of abundant rainfall, which in the higher latitudes would translate into very cold periods with ice accumulation, and hot and dry periods. Tree flora records for this period in the Iberian Peninsula are characterized by the absolute dominance of arctotercian angiosperms, with a great development of gymnosperms, among which would be Taxus, and the presence of a meager group of tropical elements, including different species of palm trees.
Dynamics of Taxus during the Pleistocene
The beginning of the Pleistocene, 2.4 Ma ago, gives way to a climate marked by the succession of cycles of glacial advance, of approximately 100,000 years duration, interspersed with phases of greater benignity, of about 10,000 years duration. In this way, during this long period there is a cyclical development of the vegetation in response to recurring climatic changes. Currently, it is recognized that this chain of climatic oscillations played a determining role in the current distribution of European tree species and their genetic patterns.
The study of Taxus dynamics during the succession of glacial cycles is fundamentally based on the analysis of pollen soundings obtained in sediments from lakes or peatlands. The data available in the European continent for the interglacials of the initial Pleistocene are reduced to few and scattered fragmented sequences. For the last glacial-interglacial cycles, the available information is expanded with the presence of continuous sequences that are still scarce and have an uneven distribution throughout the continent (Figure 6).
The paleobotanical information available for the Pleistocene reveals that Taxus has been present in Europe since the beginning of this period. In this sense, the Quaternary fossil taxon Taxus grandis, nearly a million years old, is identical in its characteristics to Taxus baccata (Hartzell, 1991). Furthermore, the pollen data obtained in various sediments attributed to the initial Pleistocene interglacials show the presence of Taxus pollen from the Initial Pleistocene, specifically from the Tiglian-C5 interglacial, 1.8 million years ago (Kase, 1988; Deforce & Bastiaens, 2007).
The pollen sequences available for the whole continental Europe and British Isles show a progressive increase in the importance of this element as the successive interglacials of the Middle Pleistocene pass, reaching its maximum landscape importance during the Holsteinian interglacial (Jessen et al., 1959, West, 1962, Kelly, 1964, Godwin, 1975, Watts, 1985, Coxon, 2017), 400–367 ky (ky: thousands of years) ago. Regarding to this interglacial period, the Taxus wooden spear recovered in 1911 (Godwin 1975, Oakley et al., 1977) in the locality of Clacton-on-Sea (Essex, United Kingdom) constitutes one of the oldest artefacts made of wood from Europe.
In the Iberian Peninsula, the oldest Pleistocene pollen remains for Taxus correspond to the record obtained in the sedimentological record of Lake Bòbila-Ordis (Leroy, 2008) located in the NE. The registry is related to an interglacial period from the early Pleistocene, although the study does not specify which of the interstate periods of the Initial Pleistocene it refers to. Other evidences of the presence of Taxus in the Iberian Peninsula correspond to the pollen grains identified in hyena coprolites recovered in the central peninsular zone and dated to the Middle Pleistocene (Carrión et al., 2006).
The last Glacial - Interglacial cycle in Northern Iberian Peninsula
Approximately 110,000 years ago, the last glacial-interglacial cycle began (Figure 5), consisting of the Eemiense interglacial (115-95 ky ago) and the Würm glacial (95-15 ky ago). The paleobotanical information available for this period in Europe increases, acquiring a better territorial representation, although it is still dispersed throughout the continent (Figure 6).
In the Iberian Peninsula, the oldest Pleistocene pollen remains for Taxus correspond to the record obtained in the sedimentological record of Lake Bòbila-Ordis (Leroy, 2008) located in the NE. The registry is related to an interglacial period from the early Pleistocene, although the study does not specify which of the interstate periods of the Initial Pleistocene it refers to. Other evidences of the presence of Taxus in the Iberian Peninsula correspond to the pollen grains identified in hyena coprolites recovered in the central peninsular zone and dated to the Middle Pleistocene (Carrión et al., 2006).
The last Glacial - Interglacial cycle in Northern Iberian Peninsula
Approximately 110,000 years ago, the last glacial-interglacial cycle began (Figure 5), consisting of the Eemiense interglacial (115-95 ky ago) and the Würm glacial (95-15 ky ago). The paleobotanical information available for this period in Europe increases, acquiring a better territorial representation, although it is still dispersed throughout the continent (Figure 6).
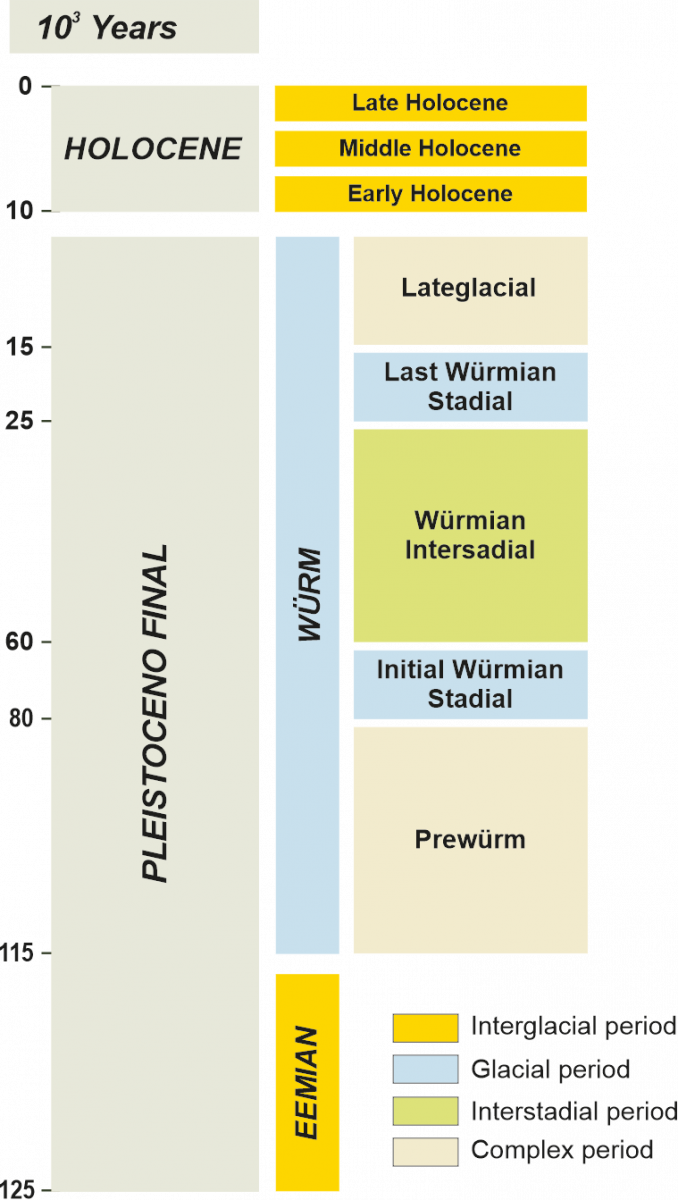
Figure 5.- Periodization of the last interglacial-glacial cycle in SW Europe.
During the last interglacial (Eemiense) the vast majority of the sequences available for the European continent in this period show an important phase of Taxus expansion, mainly in the central European territory, and decreasing its importance towards the East and the South, being even absent in some areas of the Balkan and Italic peninsulas. In the Iberian Peninsula the paleobotanical information for this interglacial is very scarce. The only data that shows the presence of Taxus is the record of some isolated pollen grains in the Eemian sequence of the Padul bog, in Granada (Camuera et al., 2019).
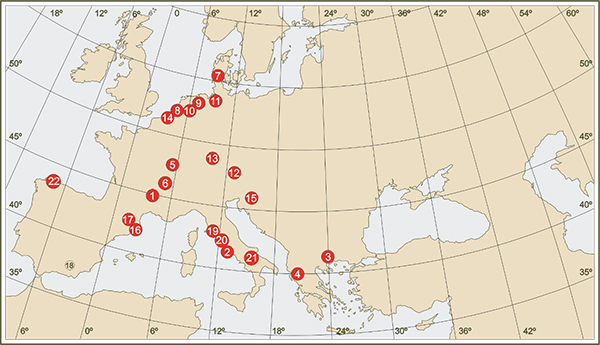
Figure 6.- Location of the main pollen sequences of the European continent that register more than one Pleistocene glacial cycle (1- Bouchet / Placraux, 2- Valle di Castiglione, 3-Tenaghi Phillipon, 4- Ioannina) or all/large part of the last glacial cycle (5- La Grande Pile, 6- Les Echets, 7- Brorup, 8- Amersfoort, 9- Denekamp, 10- Hengelo, 11- Oerel, 13- Samerberg, 14- Moershoofd, 15- Ljubljana Moor, 16 - Banyoles. 17- Pla de l'Estany, 18- Padul, 19- Lagaccione, 20- Lago di Vico, 21- Lago Grande di Monticchio, 22- Arealonga).
In the period between 95,000-15,000 years ago (Figure 5), there was a strong cooling of the climate in the Northern Hemisphere that led to important changes in the landscape and in the distribution and evolution of the wild flora and fauna lines. The mountainous territories, generally above 900-1,100 m, were affected by extremely cold conditions and dominance of ice, incompatible with the preservation of arboreal vegetation. Below the areas covered permanently or temporarily by ice, the landscape remained heavily treeless. In the Atlantic areas herbaceous formations dominated by gramineous established, while in the continental areas the lack of humidity determined the predominance of dry steppes. Below the herbaceous formations or interrelated with these in different mosaics adapted to the territory peculiarities, there were ericoid and thorny and unarmed legume shrub formations in the Atlantic areas (Heathlands), while in the continental areas Juniperus formations dominated (Juniperus communis, Juniperus sabina), as well as pulvinar legumes. The grassy and shrub formations that were dominating the landscape supported an important fauna of large, medium and small vertebrates. The forests were confined to the most protected areas of the territory, again finding a division between the Atlantic areas with formations dominated by deciduous angiosperms (Quercus, Alnus, Betula, Corylus, Castanea, Fagus, etc.) and the presence of evergreen angiosperms (Ilex, Laurus, Arbutus) and gymnosperms (Pinus, Abies, Taxus), while in the continental areas Pinus dominated and the presence of angiosperms was reduced.
In the NW of the Iberian Peninsula, paleobotanical information for the last glacial period is very limited, being limited to a series of sequences located on the current coastline. The available data do not record the presence of Taxus. The absence of pollen record from Taxus during the Würm glacial is repeated both in other areas of the Iberian Peninsula and throughout the European continent.
Taxus Dynamics during the Tardiglacial period
In the final stages of the last Glacial-Interglacial cycle, a slow increase in temperature occurred globally, which affected the configuration of the landscape and led to the current interglacial, the Holocene, extended for the last 10,000 years (Figure 7). During this period, called Tardiglacial, the ice melting that covered large terrestrial and marine surfaces happened. The domain of the ice was reduced, being in the middle of the Holocene restricted to the highest peaks. The improvement of climatic conditions determined the expansion of the forest. Tree formations were occupying more and more territory until they reached their maximum extension in the mid-Holocene, coinciding with the optimal Holocene climate. In the NW of the Iberian Peninsula the deciduous angiosperm formations became dominant in a large part of the most oceanic areas of the Cantabrian-Atlantic façade, while the interior territories were dominated by pine forests or by mosaics of pine forests, evergreen angiosperm forests and deciduous forests.
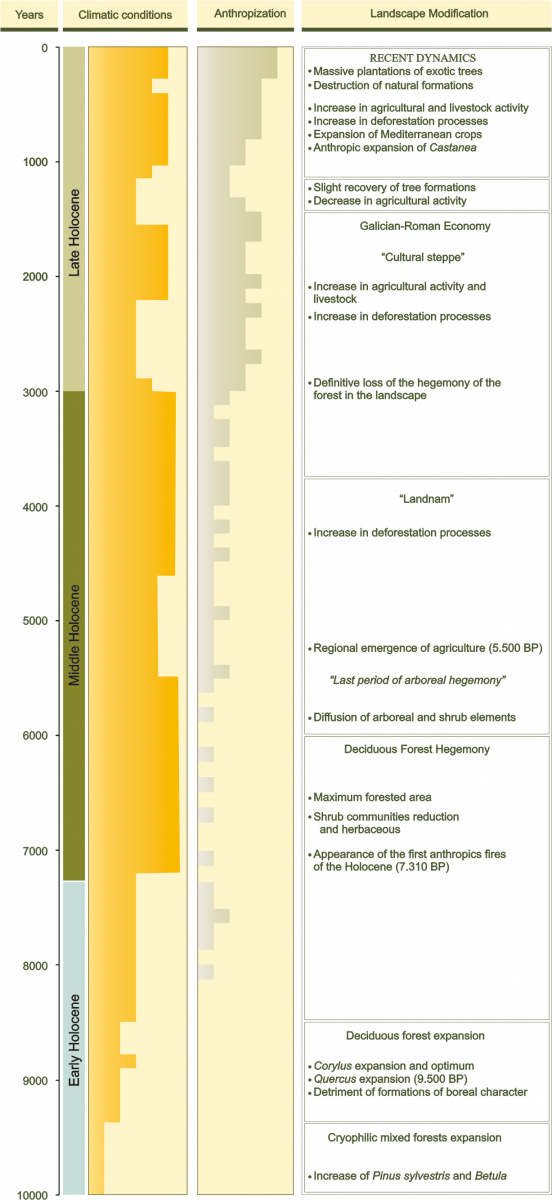
Figura 7.- Environmental periodization of Holocene in Galicia.
During the Tardiglacial deglaciation phases and the beginning of the current interglacial, the scarce presence of Taxus in the NW Iberian Peninsula diagrams shows the species persistence in the territory during the last Glacial-Interglacial cycle, occupying locations very different from the current ones, since most of these during the Pleistocene were under incompatible conditions for the species. During this thaw period, the Pena Vella sequences in the northern mountain ranges of Galicia (Ramil-Rego, 1992), La Roya in Sanabria (Allen et al., 1986), and Quintanar de la Sierra in Burgos (Peñalba, 1989), record the presence of Taxus (Table 1, Figure 8).
Dynamics of Taxus during the early Holocene phases
The yew increased its territorial distribution during the Holocene, progressing in altitude across the different regions of the Northern Iberian Peninsula (Table 1 and Figure 6). During the phases of expansion and dominance of the forest that mark the beginning of the Holocene, the number of Taxus records in NW Iberia increased. In this way, along the Cantabrian coast, various presences are registered, both from pollen grains in pollen sequences, and from coals in archaeological sites (Tables 1 and 2). In both cases, the available data refer to specific presences that would correspond to the presence of yews, as isolated individuals included within the dominant tree masses, being impossible to detect evidences of yew woods presence.
| Deposit | Tr | HI | HM | HR | Reference |
| Pena Vella | Ramil Rego, 1992 | ||||
| Pozo do Carballal | Muñoz Sobrino, 2001 | ||||
| La Roya | Allen et al., 1996 | ||||
| Cueto de Avellanosa | Núñez de la Fuente, 2018 | ||||
| Zalama | Pérez Díaz et al., 2016 | ||||
| Los Tornos | Peñalba, 1989 | ||||
| Los Tornos 3-5 | Peñalba, 1989 | ||||
| Quintanar de la Sierra | Peñalba, 1989 | ||||
| Belate | Peñalba, 1989 | ||||
| Atxurri | Peñalba, 1989 | ||||
| Saldropo 1 | Peñalba, 1989 | ||||
| Inurritza 1 | Peñalba, 1989 | ||||
| Fuente del Vaquero | Pérez-Díaz, 2012 |
Table 1.- Pollen sequences from NW Iberian Peninsula where the presence of Taxus pollen has been identified (Tl: Tardiglacial, EH: Early Holocene, MH: Medium Holocene, LH: Late Holocene).
| Deposit | HI | HM | HR | Reference |
| Atxoste | Ruíz Alonso, 2003-2007 | |||
| Buruntza | Olaetxea, 1997 | |||
| Ondarre | Mujika et al., 2013 | |||
| Mulisko Gaina | Peñalba, 1987 | |||
| Aizpea | Zapata, 2001 | |||
| Peña Larga | Zapata, 2001 | |||
| Arrubi | Ruiz-Alonso, 2003-2007 | |||
| Mendandia | Iriarte, 2006 | |||
| Peña Parda | Ruiz-Alonso & Zapata, 2003 | |||
| El Mirón | Zapata, 2012 | |||
| Mazaculos | Uzquiano, 1995 | |||
| Aramo II | Beato et al., 2019 | |||
| Peña Oviedo | Carrión Marco, 2005 | |||
| Pala da Vella | Carrión Marco, 2003 |
Table 2.- Deposits from the Iberian NW in which the presence of Taxus coal has been identified (EH: Early Holocene, MH: Medium Holocene, LH: Late Holocene).
During the Holocene initial phases, human disturbances were reduced and concentrated in certain geographical areas, intensifying and becoming generalized later in the Late Holocene, especially in territories such as Western Europe, where there was a strong reduction and isolation of natural ecosystems versus anthropoecosystems.
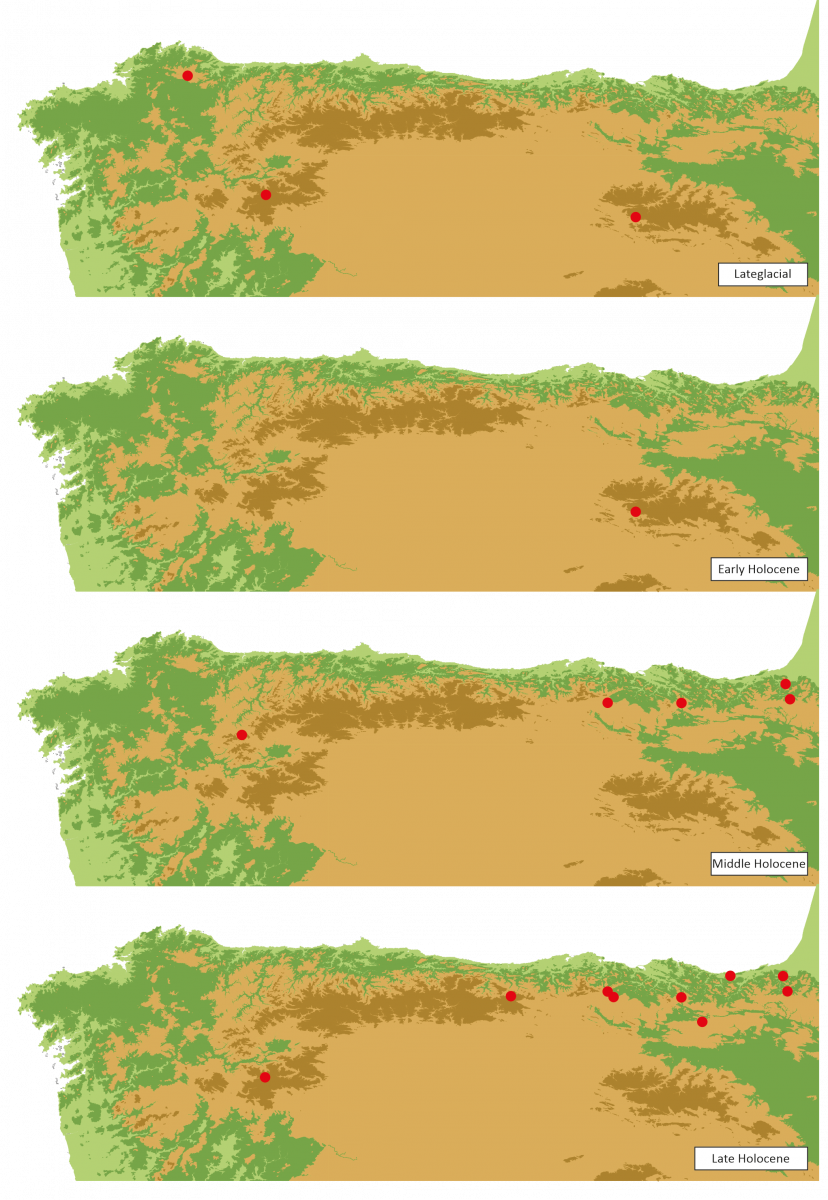
Figure 8.- Pollen sequences with presence of Taxus pollen during Tardiglacial and Holocene periods in the NW Iberian Peninsula..
Dynamics of Taxus during the last third of the Holocene
During the last third of the Holocene, pollen sequences record the appearance of Taxus pollen in some of them, while in others their record is lost (Figure 8). The registration of a greater number of localities recording Taxus pollen, with respect to Holocene previous stages, would be related to the reduction of the pollen rainfall from the dominant tree formations, which would mask in previous sequences the pollen from isolated tree elements present in those dominant forests. The reduction of the forest importance into the landscape would allow that minority elements could be more easily detected in the sequences.
The adoption of crop and livestock production systems brought fundamental changes in the configuration of the landscape throughout Europe. In the Northern Iberian Peninsula this process began 5,500 years ago when the first records of cultivated plants were recorded. Coinciding with the end of the Bronze Age and the beginning of the Iron Age, the small villages of farmers and ranchers increase their production and therefore their footprint on the landscape. So that 2,000 years ago in the most intensely populated areas, forests and natural vegetation are displaced by the configuration of agrosystems supported by the force of people and domestic animals.
During this period yew is scarce in the landscape, and like other tree species, it is sheltered in mountain areas where agricultural and livestock pressures were less important. The open landscapes configured by agrosystems were maintained and reinforced during the Middle Ages and Modern Ages, increasing the confinement of woodland to mountain areas. In many sites, especially in the mountains, the yew trees acquired a special role during this period as a medicinal, symbolic and spiritual resource, integrating itself into domestic and religious spaces where they remain as living witnesses of a complex and remote past, while the native formations faded and finally disappeared.
Yew formations and forests are a basic component of biodiversity in Europe, forged over thousands of years as a result of major biogeographic and climatic changes. Forests and formations that have maintained and still maintain in many areas, as in the Atlantic Region of the Iberian Peninsula, an intricate relationship with local communities that have benefited from the ecosystem services they report.
References
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Beato Bergua, S., Poblete Piedrabuena, M.A., Cunill Artigas, R. (2019). Taxus baccata en la Sierra del Aramo (Macizo Central Asturiano). Boletín de la Asociación de Geógrafos Españoles, 81, 2772, 1–30.
Camuera, J., Jiménez-Moreno, G., Ramos-Román, M.J., García-Alix, A., Toney, J.L., Anderson, R.S., Jiménez-Espejo, F., Bright, J., Webster, C., Yanes, Y., Carrión, J.S. (2019). Vegetation and climate changes during the last two glacial-interglacial cycles in the western Mediterranean: A new long pollen record from Padul (southern Iberian Peninsula),Quaternary Science Reviews, 205: 86-105.
Carrión Marco, Y. (2003). El paisaje vegetal prehistórico de Pala da Vella: primeros resultados antracológicos. In: V. Oliveira Jorge. Contributos das ciencias e das tecnologias para a arqueologia da Península Ibérica: 21-32. UTAD Vila Real, Portugal
Carrión Marco, Y. (2005). El impacto de la economía productora en el paisaje vegetal de Peña Oviedo (Camaleño, Cantabria). In: R. Ontañón Peredo, C. García-Moncó Piñeiro, P. Arias Cabal. Actas del III Congreso del Neolítico en la Península Ibérica: 35-44. Santander.
Carrión, J. S., Scott, L., Arribas, A., Fuentes, N., Gil‐Romera, G. and Montoya, E. (2006). Pleistocene landscapes in central Iberia inferred from pollen analysis of hyena coprolites. Journal of Quaternary Science, 22: 191–202.
Coxon, P., McCarron, S., Mitchell, F. (Eds) (2017). Advances in Irish Quaternary Studies. Atlantis Advances in Quaternary Science. Atlantis Press, Paris. 316pp.
Deforce, K. & Bastiaens, J. (2007). The Holocene history of Taxus baccata (yew) in Belgium and neighbouring regions. Belgian Journal of Botany, 140(2): 222-237.
Florin, R. (1958). On Jurassic taxads and conifers from North-Western Europe and Eastern Greenland. Acta Horti Bergiani Series, 17: 257-402.
Godwin H. (1975). The history of the British Flora. A factual basis for phytogeography. Cambridge University Press, Cambridge. 383 pp.
Hao, D.-C., Huang, B., Yang, L. (2008). Phylogenetic Relationships of the Genus Taxus Inferred from Chloroplast Intergenic Spacer and Nuclear Coding DNA. Biological & pharmaceutical bulletin, 31(2): 260-265.
Hartzell, J.R. (1991). The Yew Tree: A Thousand Whispers. Hulogosi Communications, Oregon, 319 pp.
Heim J. (1970). Les relations entre les spectres polliniques récents et la végétation actuelle en Europe occidentale. Ph.D. thesis, Université catholique de Louvain, Louvain-la-Neuve, Belgium.
Iriarte, M.J. (2006). El entorno vegetal del abrigo de Mendandia y su depósito arqueológico: Análisis palinológico. In: Alday A. (Dir.). El legado arqueologico de Mendandia. Los modos de vida de los últimos cazadores en la Prehistoria de Treviño: 405-418. Junta de Castilla y León. Burgos.
Jessen, K., Andersen, S.Th., Farrington, A. (1959). The interglacial deposit near Gort, Co. Galway, Ireland. Proceedings of the Royal Irish Academy B, 60: 1-77.
Kasse K. (1988). Early-Pleistocene tidal and fluviatile environments in the southern Netherlands and northern Belgium. Ph.D. thesis, Vrije Universiteit, Amsterdam, The Netherlands.
Kelly, M.R. (1964) The Middle Pleistocene of North Birmingham. Philos. T. Roy. Soc. B, 247: 533-592.
Kunzmann L., Mai D.H. (2005) Die Koniferen der Mastixioideen-Flora von Wiesa bei Kamenz (Sachsen, Miozän) unter besonderer Berücksichtigung der Nadelblätter. Palaeontographica B 272(1–6): 67–135.
Leroy, S.A.G. (2008): Vegetation cycles in a disturbed sequence around the Cobb-Mountain subchron in Catalonia (Spain). Journal of Paleolimnology, 40(3), 851-868
Li, J., Davis, C.C., Del Tredici, P., Donoghue, M.J. (2001). Phylogeny and biogeography of Taxus (Taxaceae) inferred from sequences of the internal transcribed spacer region of nuclear ribosomal DNA. Harv. Pap. Bot. ,6:267–274.
Mujika-Alustiza, J.A., Agirre-García, J., Edeso-Fito, J.M., Lopetegi-Galarraga, A., Pérez-Díaz, S., Ruiz-Alonso, M., Tarriño-Vinagre, A., Yusta-Arnal, I. (2013). La continuidad de la actividad pastoril durante la época romana en la zona de Argarbi (sierra de Aralar - Gipuzkoa). Kobie Serie PaleoantroPología, 32: 217-258
Muñoz Sobrino, C. (2001). Cambio climático y dinámica del paisaje en las montañas del noroeste de la Península Ibérica. Tesis Doctoral. Escuela Politécnica Superior. Universidad de Santiago de Compostela.
Nathorst A.G. (1908). Palaeobotanische Mitteillungen.7. Über Palyssya, Stachyotaxus und Paleotaxus. Ksvenka Vetensk Akad Handl 8: 1–16.
Nuñez de la Fuente, S. (2018). Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje atlántico montañoso: la región cantábrica durante el Holoceno. Tesis doctoral. Universidad de Cantabria.
Peñalba Garmendia, M.C. (1989). Dynamique de Végétation Tardiglaciaire et Holocene Du Centre-Nord de l’Espagne d’après l’analyse pollinique. These. Université d’Aix, Marseille III.
Pérez-Díaz, S. (2012). El paisaje vegetal durante la Prehistoria Reciente en la vertiente mediterránea de Euskal Herria— Tesis doctoral. Euskal Herriko Univ., Vitoria-Gastei.
Perez-Díaz, S., López-Sáez, J. A., Pontevedra-Pombal, X., Souto-Souto, M. & Galop, D. (2016). 8000 years of vegetation history in the northern Iberian Peninsula inferred from the palaeoenvironmental study of the Zalama ombrotrophic bog (Basque-Cantabrian Mountains, Spain). Boreas, Vol. 45, pp. 658–672.
Ramil Rego P, Gómez-Orellana L, Muñoz Sobrino C, Tereso JP (2011). Dinâmica natural e transformação antrópica das florestas do noroeste ibérico. In Tereso JP, Honrado JP, Pinto AT, Rego FC (Eds.) Florestas do Norte de Portugal: História, Ecologia e Desafios de Gestão. InBio - Rede de Investigação em Biodiversidade e Biologia Evolutiva. Porto. ISBN: 978-989-97418-1-2. Pp 14-54.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ruiz-Alonso, M. & Zapata, L. (2003). Análisis antracológico del yacimiento arqueológico de Peña Parda. Cuad. Arq. Univ. Navarra, 11: 217-252.
Ruiz-Alonso, M. (2003/2007). Madera carbonizada en los fondos de cabaña de Arrubi y Esnaurreta (Sierra de Aralar, Gipuzkoa): vegetación y recursos forestales en la Edad Media. Kobie (Serie Paleoantr.), 27: 131-150.
Uzquiano, P. (1995). L’evolution de la vegetation a l’Holocene inicial dans le nord de l’Espagne a partir de l’étude anthracologique de trois sites archeologique. Quaternaire 6 (2): 77-83.
Van der Knaap, W.O., van Leeuwen, J.F.N. (1997). Late Glacial and early Holocene vegetation succession, altitudinal vegetation zonation, and climatic change in the Serra da Estrela, Portugal. Review of Palaeobotany and Palynology, 97:3-4: 239-285.
Watts, W.A. (1985) Quaternary vegetation cycles.In: Edwards, K.J. & Warren, W.P. (eds.): The Quaternary history of Ireland: 154-185. Academic Press, London.
West, R.G. (1962) A note on Taxus pollen in the Hoxnian Interglacial. New Phytologist, 61: 189-190.
Zapata, L. (2012). Holocene wood charcoal from El Mirón cave. Vegetation and wood use — In: Straus, L.G. & González Morales, M.R. (Eds.). El Miron cave. Cantabrian Spain. The site and its Holocene archaeological record: 174-196. Univ. New Mexico, Alburquerque.
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Beato Bergua, S., Poblete Piedrabuena, M.A., Cunill Artigas, R. (2019). Taxus baccata en la Sierra del Aramo (Macizo Central Asturiano). Boletín de la Asociación de Geógrafos Españoles, 81, 2772, 1–30.
Camuera, J., Jiménez-Moreno, G., Ramos-Román, M.J., García-Alix, A., Toney, J.L., Anderson, R.S., Jiménez-Espejo, F., Bright, J., Webster, C., Yanes, Y., Carrión, J.S. (2019). Vegetation and climate changes during the last two glacial-interglacial cycles in the western Mediterranean: A new long pollen record from Padul (southern Iberian Peninsula),Quaternary Science Reviews, 205: 86-105.
Carrión Marco, Y. (2003). El paisaje vegetal prehistórico de Pala da Vella: primeros resultados antracológicos. In: V. Oliveira Jorge. Contributos das ciencias e das tecnologias para a arqueologia da Península Ibérica: 21-32. UTAD Vila Real, Portugal
Carrión Marco, Y. (2005). El impacto de la economía productora en el paisaje vegetal de Peña Oviedo (Camaleño, Cantabria). In: R. Ontañón Peredo, C. García-Moncó Piñeiro, P. Arias Cabal. Actas del III Congreso del Neolítico en la Península Ibérica: 35-44. Santander.
Carrión, J. S., Scott, L., Arribas, A., Fuentes, N., Gil‐Romera, G. and Montoya, E. (2006). Pleistocene landscapes in central Iberia inferred from pollen analysis of hyena coprolites. Journal of Quaternary Science, 22: 191–202.
Coxon, P., McCarron, S., Mitchell, F. (Eds) (2017). Advances in Irish Quaternary Studies. Atlantis Advances in Quaternary Science. Atlantis Press, Paris. 316pp.
Deforce, K. & Bastiaens, J. (2007). The Holocene history of Taxus baccata (yew) in Belgium and neighbouring regions. Belgian Journal of Botany, 140(2): 222-237.
Florin, R. (1958). On Jurassic taxads and conifers from North-Western Europe and Eastern Greenland. Acta Horti Bergiani Series, 17: 257-402.
Godwin H. (1975). The history of the British Flora. A factual basis for phytogeography. Cambridge University Press, Cambridge. 383 pp.
Hao, D.-C., Huang, B., Yang, L. (2008). Phylogenetic Relationships of the Genus Taxus Inferred from Chloroplast Intergenic Spacer and Nuclear Coding DNA. Biological & pharmaceutical bulletin, 31(2): 260-265.
Hartzell, J.R. (1991). The Yew Tree: A Thousand Whispers. Hulogosi Communications, Oregon, 319 pp.
Heim J. (1970). Les relations entre les spectres polliniques récents et la végétation actuelle en Europe occidentale. Ph.D. thesis, Université catholique de Louvain, Louvain-la-Neuve, Belgium.
Iriarte, M.J. (2006). El entorno vegetal del abrigo de Mendandia y su depósito arqueológico: Análisis palinológico. In: Alday A. (Dir.). El legado arqueologico de Mendandia. Los modos de vida de los últimos cazadores en la Prehistoria de Treviño: 405-418. Junta de Castilla y León. Burgos.
Jessen, K., Andersen, S.Th., Farrington, A. (1959). The interglacial deposit near Gort, Co. Galway, Ireland. Proceedings of the Royal Irish Academy B, 60: 1-77.
Kasse K. (1988). Early-Pleistocene tidal and fluviatile environments in the southern Netherlands and northern Belgium. Ph.D. thesis, Vrije Universiteit, Amsterdam, The Netherlands.
Kelly, M.R. (1964) The Middle Pleistocene of North Birmingham. Philos. T. Roy. Soc. B, 247: 533-592.
Kunzmann L., Mai D.H. (2005) Die Koniferen der Mastixioideen-Flora von Wiesa bei Kamenz (Sachsen, Miozän) unter besonderer Berücksichtigung der Nadelblätter. Palaeontographica B 272(1–6): 67–135.
Leroy, S.A.G. (2008): Vegetation cycles in a disturbed sequence around the Cobb-Mountain subchron in Catalonia (Spain). Journal of Paleolimnology, 40(3), 851-868
Li, J., Davis, C.C., Del Tredici, P., Donoghue, M.J. (2001). Phylogeny and biogeography of Taxus (Taxaceae) inferred from sequences of the internal transcribed spacer region of nuclear ribosomal DNA. Harv. Pap. Bot. ,6:267–274.
Mujika-Alustiza, J.A., Agirre-García, J., Edeso-Fito, J.M., Lopetegi-Galarraga, A., Pérez-Díaz, S., Ruiz-Alonso, M., Tarriño-Vinagre, A., Yusta-Arnal, I. (2013). La continuidad de la actividad pastoril durante la época romana en la zona de Argarbi (sierra de Aralar - Gipuzkoa). Kobie Serie PaleoantroPología, 32: 217-258
Muñoz Sobrino, C. (2001). Cambio climático y dinámica del paisaje en las montañas del noroeste de la Península Ibérica. Tesis Doctoral. Escuela Politécnica Superior. Universidad de Santiago de Compostela.
Nathorst A.G. (1908). Palaeobotanische Mitteillungen.7. Über Palyssya, Stachyotaxus und Paleotaxus. Ksvenka Vetensk Akad Handl 8: 1–16.
Nuñez de la Fuente, S. (2018). Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje atlántico montañoso: la región cantábrica durante el Holoceno. Tesis doctoral. Universidad de Cantabria.
Peñalba Garmendia, M.C. (1989). Dynamique de Végétation Tardiglaciaire et Holocene Du Centre-Nord de l’Espagne d’après l’analyse pollinique. These. Université d’Aix, Marseille III.
Pérez-Díaz, S. (2012). El paisaje vegetal durante la Prehistoria Reciente en la vertiente mediterránea de Euskal Herria— Tesis doctoral. Euskal Herriko Univ., Vitoria-Gastei.
Perez-Díaz, S., López-Sáez, J. A., Pontevedra-Pombal, X., Souto-Souto, M. & Galop, D. (2016). 8000 years of vegetation history in the northern Iberian Peninsula inferred from the palaeoenvironmental study of the Zalama ombrotrophic bog (Basque-Cantabrian Mountains, Spain). Boreas, Vol. 45, pp. 658–672.
Ramil Rego P, Gómez-Orellana L, Muñoz Sobrino C, Tereso JP (2011). Dinâmica natural e transformação antrópica das florestas do noroeste ibérico. In Tereso JP, Honrado JP, Pinto AT, Rego FC (Eds.) Florestas do Norte de Portugal: História, Ecologia e Desafios de Gestão. InBio - Rede de Investigação em Biodiversidade e Biologia Evolutiva. Porto. ISBN: 978-989-97418-1-2. Pp 14-54.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ruiz-Alonso, M. & Zapata, L. (2003). Análisis antracológico del yacimiento arqueológico de Peña Parda. Cuad. Arq. Univ. Navarra, 11: 217-252.
Ruiz-Alonso, M. (2003/2007). Madera carbonizada en los fondos de cabaña de Arrubi y Esnaurreta (Sierra de Aralar, Gipuzkoa): vegetación y recursos forestales en la Edad Media. Kobie (Serie Paleoantr.), 27: 131-150.
Uzquiano, P. (1995). L’evolution de la vegetation a l’Holocene inicial dans le nord de l’Espagne a partir de l’étude anthracologique de trois sites archeologique. Quaternaire 6 (2): 77-83.
Van der Knaap, W.O., van Leeuwen, J.F.N. (1997). Late Glacial and early Holocene vegetation succession, altitudinal vegetation zonation, and climatic change in the Serra da Estrela, Portugal. Review of Palaeobotany and Palynology, 97:3-4: 239-285.
Watts, W.A. (1985) Quaternary vegetation cycles.In: Edwards, K.J. & Warren, W.P. (eds.): The Quaternary history of Ireland: 154-185. Academic Press, London.
West, R.G. (1962) A note on Taxus pollen in the Hoxnian Interglacial. New Phytologist, 61: 189-190.
Zapata, L. (2012). Holocene wood charcoal from El Mirón cave. Vegetation and wood use — In: Straus, L.G. & González Morales, M.R. (Eds.). El Miron cave. Cantabrian Spain. The site and its Holocene archaeological record: 174-196. Univ. New Mexico, Alburquerque.