
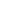


Historia de los bosques de tejo en el Norte de la Península Ibérica
11-12-2020
La tierra a lo largo del tiempo ha sufrido numerosas transformaciones que condicionaron la vida sobre el planeta. Las principales alteraciones en la historia de la tierra, fueron la deriva continental y el cambio climático. La fuerza más importante del cambio climático en el pasado era la modificación de los ciclos orbitales de la tierra, que en último término determinan la cantidad de insolación que recibe cada parte del planeta y por tanto el clima en las distintas zonas del globo. Sin embargo, los últimos años de la historia de la tierra están marcados por las modificaciones derivadas de la alteración humana en los ecosistemas naturales.
La información paleoecológica nos permite conocer la evolución del clima y su influencia sobre los diferentes biomas y especies. En este sentido, el estudio de los restos vegetales proporciona una de las visiones más precisas de la dinámica de los ecosistemas y las especies. Los análisis paleobotánicos permiten analizar la respuesta de las diferentes especies frente a los cambios climáticos u otras perturbaciones.
A partir del estudio de los macrofósiles (restos de troncos, ramas, hojas, frutos, etc.) y microfósiles (granos de polen) de las especies arbóreas, puede reconstruirse su evolución en un territorio determinado, observando como las diferentes alteraciones naturales y antrópicas influyeron en su distribución a lo largo del tiempo y evaluando los fenómenos de migración, refugio, extinción, etc.
Aparición de las Taxaceae
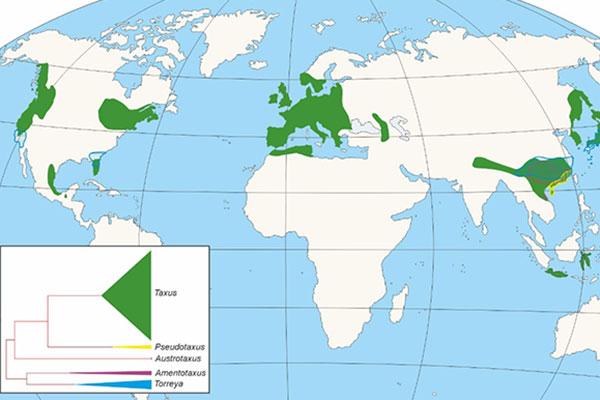
Dentro de las gimnospermas, la familia Taxaceae constituye un grupo claramente diferenciado del resto de las coníferas, estando representado en la actualidad por cinco géneros divididos evolutivamente en dos clados, uno con tres géneros Taxus, Austrotaxus y Pseudotaxus, y el segundo con dos (Torreya y Amentotaxus), que en conjunto reúnen unas 28 especies distribuidas en Eurasia, Norte de África, Nueva Caledonia y América (Figura 1), a las que hay que unir distintas especies extintas.
La información paleoecológica nos permite conocer la evolución del clima y su influencia sobre los diferentes biomas y especies. En este sentido, el estudio de los restos vegetales proporciona una de las visiones más precisas de la dinámica de los ecosistemas y las especies. Los análisis paleobotánicos permiten analizar la respuesta de las diferentes especies frente a los cambios climáticos u otras perturbaciones.
A partir del estudio de los macrofósiles (restos de troncos, ramas, hojas, frutos, etc.) y microfósiles (granos de polen) de las especies arbóreas, puede reconstruirse su evolución en un territorio determinado, observando como las diferentes alteraciones naturales y antrópicas influyeron en su distribución a lo largo del tiempo y evaluando los fenómenos de migración, refugio, extinción, etc.
Aparición de las Taxaceae
Dentro de las gimnospermas, la familia Taxaceae constituye un grupo claramente diferenciado del resto de las coníferas, estando representado en la actualidad por cinco géneros divididos evolutivamente en dos clados, uno con tres géneros Taxus, Austrotaxus y Pseudotaxus, y el segundo con dos (Torreya y Amentotaxus), que en conjunto reúnen unas 28 especies distribuidas en Eurasia, Norte de África, Nueva Caledonia y América (Figura 1), a las que hay que unir distintas especies extintas.
Figura 1. Distribución de los géneros de la Familia Taxaceae y cladograma mostrando la aparición e importancia de cada uno de ellos.
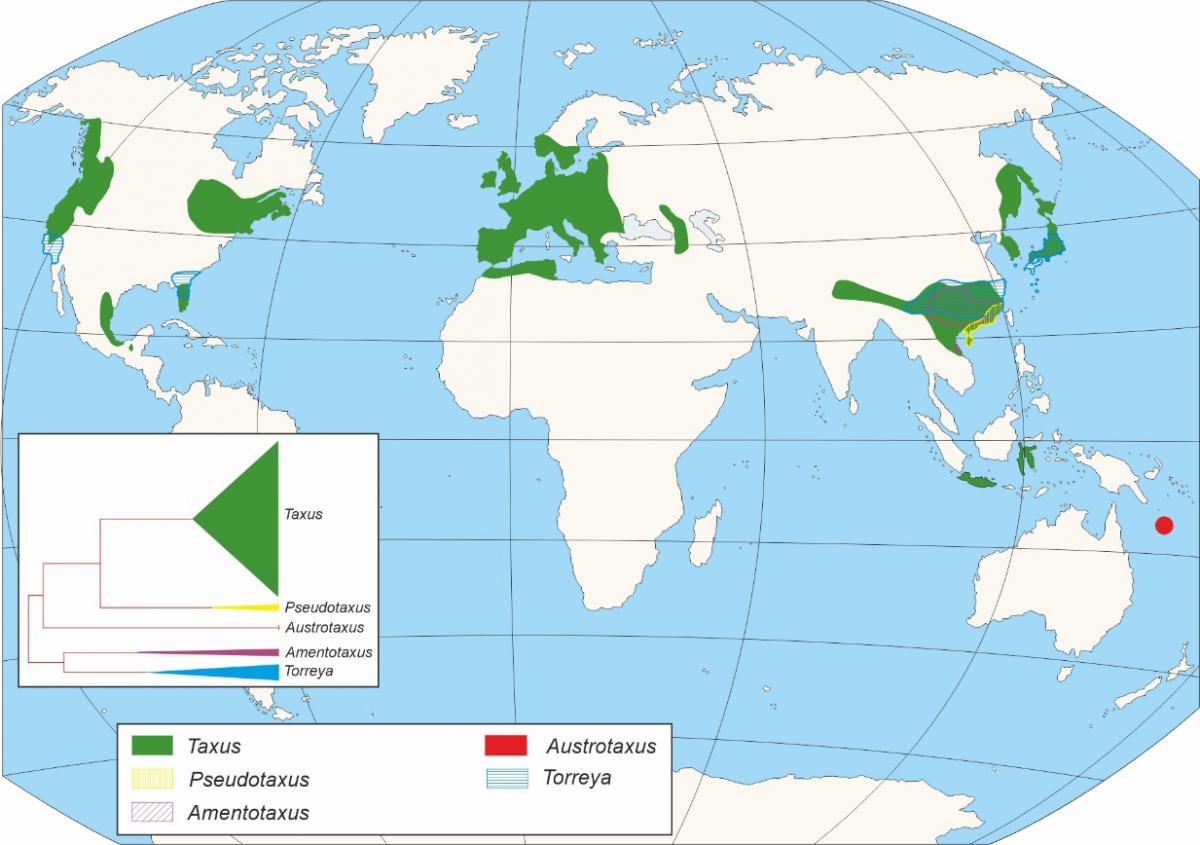
La aparición de la familia de las Taxaceae se sitúa en el Mesozoico (Figura 2), concretamente en el periodo Triásico, hace unos 250-200 Ma (Ma: millones de años). Taxaceae irradió desde el suroeste de China, divergiendo en varios clados que incluirían el linaje europeo (Figura 1). El fósil de mayor antigüedad de Taxaceae, se corresponde con un fragmento de una rama con hojas de Paleotaxus rediviva recuperado en una facies de 200 Ma de antigüedad en 1908 en el sur de Suecia (Nathorst 1908). A partir de Paleotaxus rediviva, evolucionaria durante el Jurásico, hace 140 Ma, el grupo Taxus jurassica (Figura 2). Posteriormente, durante el Cretácico y Cenozoico, se diferencian nuevas especies, en un proceso que estuvo modulado por importantes cambios tanto en la delimitación de las unidades continentales, como en el clima, que afectaron a su distribución y al aislamiento de distintos grupos poblacionales en distintos territorios del hemisferio Norte.
Figura 2.- Localización en la escala geológica de los diferentes eventos relacionados con la aparición de Taxus. Además, se muestran otros eventos significa
tivos. Ma: millones de años.
El género Taxus (Figuras 2 y 3) está conformado por árboles dioicas con ramificación irregular, con hojas aciculares y yemas escamadas siendo el de mayor importancia de la familia Taxaceae. El género Taxus está integrado por al menos diez especies, aunque existe un debate en cuanto al número de las mismas, principalmente en el continente asiático, considerándose una cuestión aun no resuelta (Hao et al., 2008). Los primeros restos fósiles adscritos al género Taxus, fueron designados como Taxus jurassica (140 Ma). Las características morfológicas de estos permiten su asimilación con distintas especies actuales: T. baccata, T. cuspidata, T. brevifolia (Florin, 1958).
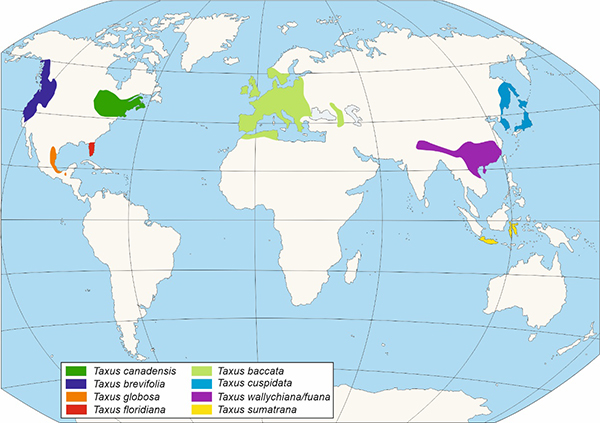
Figura 3. – Distribución de las distintas especies del Género Taxus según Li et al., 2001.
La era Mesozoica (Figura 2) se extiende entre hace 245 Ma, hasta hace 65 Ma. Durante la misma, la masa continental que constituía el supercontinente Pangea permanece estable sin sufrir grandes alteraciones. Las condiciones climáticas permanecieron básicamente estables, predominando a lo largo del continente los ambientes tropicales y subtropicales que marcarían el predominio de los taxa de distribución cosmopolita. Las plantas con flores dominantes durante esta era son las gimnospermas, entre las cuales se incluirían Taxaceae y otras coníferas (como los ginkgos y las cycas), mientras que, en la parte final del Mesozoico, a partir del cretácico inferior, comienzan a aparecer los primeros fósiles de angiospermas (Figura 2).
Teniendo en cuenta los datos paleobotánicos, Taxus es el género de árboles más antiguo del continente europeo, considerándose un género relicto. El fósil de Taxus de mayor antigüedad, recuperado en Europa, se sitúa en los inicios del Mioceno, hace unos 23 Ma (Figura 2). Se corresponde con unas hojas de Taxus engelhardtii localizadas en la localidad de Wiesa en Sajonia (Kunzmann & Mai, 2005). El inicio del Mioceno determina el inicio de una larga etapa de inestabilidad climática a nivel planetario, que marcará la reducción de los bosques y especies tropicales dominantes y el paulatino incremento de importancia de los elementos arctoterciarios y esclerófilos. En relación con la transición entre Mioceno y Plioceno, se han recuperado diferentes macrofósiles, incluyendo semillas, ramitas u hojas, en localidades dispersas del Centro y Este de Europa.
En la Península Ibérica, los restos fósiles más antiguos se corresponden con granos de polen recuperados en diversas secuencias obtenidas dentro de la cuenca lacustre Plio-pleistocena de Banyoles-Besalú, en Girona (Leroy, 2008) y en las cuencas mineras de Galicia, Norte y Centro de Portugal (Ramil-Rego et al., 2011). En estos entornos deposicionales, se registran algunos granos de polen aislados de Taxus o Taxaceae, relacionados con la época Pliocena (hace 5 millones de años)
 Figura 4.- Tronco fósil de gimnosperma (aprox. 150 millones de años), recogida por el Profesor Montenegro de Andrade en Caldas da Rainha (Portugal) y expuesto en el edificio del rectorado de la
Figura 4.- Tronco fósil de gimnosperma (aprox. 150 millones de años), recogida por el Profesor Montenegro de Andrade en Caldas da Rainha (Portugal) y expuesto en el edificio del rectorado de laUniversidade do Porto.
El clima plioceno, se caracteriza por la alternancia de periodos fríos de abundantes lluvias, que en las latitudes más altas se traducirían en periodos muy fríos con acumulación de hielo, y periodos secos y cálidos. Los registros de flora arbórea para este periodo en la Península Ibérica se caracterizan por el dominio absoluto de las angiospermas arctoterciarias, con un gran desarrollo de las gimnospermas, entre las que se encontraría Taxus, y la presencia de un exiguo grupo de elementos tropicales, entre los que se incluyen distintas especies de palmeras.
Dinámica de Taxus durante el Pleistoceno
El inicio del Pleistoceno, hace 2,4 Ma, da paso a un clima marcado por la sucesión de ciclos de avance glaciar, de aproximadamente 100.000 años de duración, intercalados con fases de mayor benignidad, de unos 10.000 años de duración. De este modo, durante este dilatado periodo se produce un desarrollo cíclico de la vegetación en respuesta a los cambios climáticos de carácter recurrente. Actualmente, se reconoce que este encadenamiento de oscilaciones climáticas jugaron un papel determinante en la actual distribución de las especies arbóreas europeas y de sus patrones genéticos.
El estudio de la dinámica de Taxus durante la sucesión de ciclos glaciares se basa fundamentalmente en el análisis de sondeos polínicos obtenidos en sedimentos de lagos o turberas. Los datos disponibles en el continente europeo para los interglaciares del Pleistoceno inicial se reducen a escasas y dispersas secuencias fragmentadas. Para los últimos ciclos glaciar-interglaciar la información disponible se amplía con la presencia de secuencias continuas que aún son escasas y presentan una desigual distribución a lo largo del continente (Figura 6).
La información paleobotánica disponible para el Pleistoceno revela que Taxus está presente en Europa desde el inicio de este periodo. En este sentido, el taxón fósil cuaternario Taxus grandis, con cerca de un millón de años de antigüedad, es idéntico en sus características a Taxus baccata (Hartzell, 1991). Además, los datos polínicos obtenidos en diversos sedimentos atribuidos a las interglaciares iniciales del Pleistoceno muestran la presencia de polen de Taxus desde el Pleistoceno Inicial, concretamente desde el interglaciar Tiglian-C5, hace 1,8 millones de años, (Kase, 1988; Deforce & Bastiaens, 2007).
Las secuencias polínicas disponibles para el conjunto de Europa continental y las islas británicas muestran un progresivo incremento de la importancia de este elemento a medida que transcurren los sucesivos interglaciares del Pleistoceno medio, alcanzando su máxima importancia paisajística durante el interglaciar Holsteinian (Jessen et al., 1959, West, 1962, Kelly, 1964, Godwin, 1975, Watts, 1985, Coxon, 2017), hace 400–367 ka (ka: miles de años). En relación con este interglaciar se sitúa la lanza de madera de Taxus recuperada en 1911 (Godwin 1975, Oakley et al., 1977) en la localidad de Clacton-on-Sea (Essex, Reino Unido), que constituye uno de los más antiguos artefactos fabricados en madera de Europa.
En la Península Ibérica, los restos polínicos pleistocenos de mayor antigüedad para Taxus se corresponden con el registro obtenido en el registro sedimentológico del lago de Bòbila-Ordis (Leroy, 2008) situado en el NE. El registro se relaciona con un periodo interglaciar de los inicios del Pleistoceno, aunque en el estudio no se precisa a cuál de los periodos interestadiales del Pleistoceno Inicial se refiere. Otras evidencias de la presencia de Taxus en la Península Ibérica se corresponden con los granos de polen identificados en coprolitos de hiena recuperados en la zona central peninsular y datados para el Pleistoceno Medio (Carrión et al., 2006).
El último ciclo Glaciar – Interglaciar en el Norte de la Península Ibérica
Hace aproximadamente 110.000 años se inicia el último ciclo glaciar-interglaciar (Figura 5), integrado por el interglaciar Eemiense (hace 115-95 ka) y el glaciar Würm (hace 95-15 ka). La información paleobotánica disponible para este periodo en Europa se incrementa, adquiriendo una mejor representación territorial, aunque aún dispersa a lo largo del continente (Figura 6).
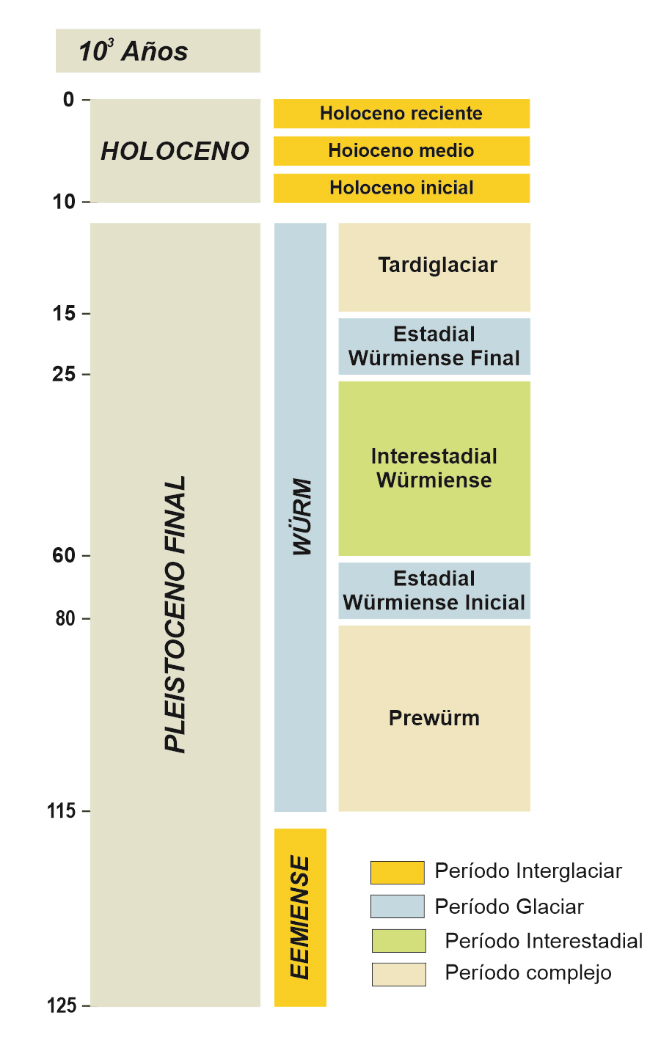
Figura 5.- Periodización del último ciclo glaciar interglaciar en el SW de Europa.
Durante el último interglaciar (Eemiense) la gran mayoría de las secuencias disponibles para el continente europeo en este periodo muestran una importante fase de expansión de Taxus, principalmente en el territorio centro europeo, y disminuyendo su importancia hacia el Este y el Sur, estando incluso ausente en algunas áreas de las penínsulas balcánica e itálica. En la Península Ibérica la información paleobotánica para este interglaciar es muy escasa. El único dato que muestra la presencia de Taxus es el registro de unos granos de polen aislados en la secuencia Eemiense de la turbera de Padul, en Granada (Camuera et al., 2019).
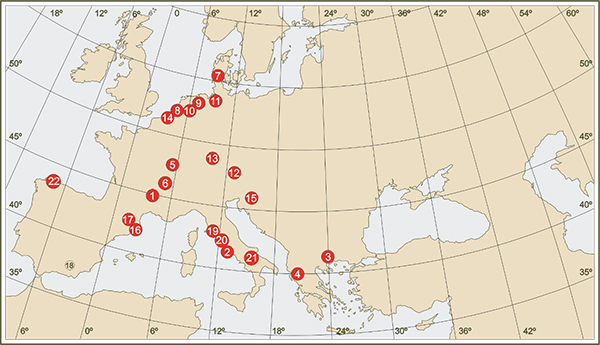
Figura 6.- Localización de las principales secuencias polínicas del continente europeo que registran más de un ciclo glaciar del pleistoceno (1- Bouchet/Placraux, 2- Valle di Castiglione, 3-Tenaghi Phillipon, 4- Ioannina) o la totalidad o gran parte del último ciclo glaciar (5- La Grande Pile, 6- Les Echets, 7- Brorup, 8- Amersfoort, 9- Denekamp, 10- Hengelo, 11- Oerel, 13- Samerberg, 14- Moershoofd, 15- Ljubljana Moor, 16- Banyoles. 17- Pla de l’Estany, 18- Padul, 19- Lagaccione, 20- Lago di Vico, 21- Lago Grande di Monticchio, 22- Arealonga).
En el periodo comprendido entre hace 95.000-15.000 años (Figura 5) se produjo en el hemisferio Norte un fuerte enfriamiento del clima que condujo a importantes cambios en el paisaje y en la distribución y evolución de las estirpes de flora y fauna silvestre. Los territorios montañosos, generalmente por encima de los 900-1.100 m, estuvieron afectados por condiciones extremadamente frías y dominio de los hielos, incompatibles para la preservación de la vegetación arbórea. Por debajo de las áreas cubiertas de forma permanente o temporal por los hielos el paisaje se mantenía fuertemente desarbolado. En las áreas atlánticas se establecen formaciones herbosas dominadas por gramíneas, mientras que en las continentales la falta de humedad determina el predominio de estepas secas. Por debajo de las formaciones herbáceas o interrelacionadas con estas en distintos mosaicos adaptados a las peculiaridades del territorio, se encuentran formaciones arbustivas, de ericoides y leguminosas espinosas e inermes en las áreas Atlánticas (Brezales), mientras que en las áreas continentales dominan formaciones de Juniperus (Juniperus communis, Juniperus sabina), así como de leguminosas pulvinulares. Las formaciones herbosas y arbustivas que dominan el paisaje sustentan una importante fauna de grandes, medianos y pequeños vertebrados. Los bosques quedan confinados a las áreas más protegidas del territorio, encontrando de nuevo una división entre las áreas Atlánticas con formaciones dominadas por angiospermas caducifolias (Quercus, Alnus, Betula, Corylus, Castanea, Fagus, etc.) y presencia de perennifolias (Ilex, Laurus, Arbutus) y de gimnospermas (Pinus, Abies, Taxus), mientras que en las áreas continentales dominan los Pinus y se reduce la presencia de angiospermas.
En el NW de la Península Ibérica, la información paleobotánica para el último periodo glaciar es muy reducida, limitándose a una serie de secuencias situadas en la actual línea de costa. Los datos disponibles no registran la presencia de Taxus. La ausencia de registro de polen de Taxus durante el glaciar Würm se repite tanto en otras áreas de la Península Ibérica como a lo largo del continente europeo.
Dinámica de Taxus durante el Tardiglaciar
En las etapas finales del último ciclo Glaciar-Interglaciar se produce globalmente un lento aumento de la temperatura que repercute en la configuración del paisaje y que desemboca en el actual interglaciar, el Holoceno, que se extiende entre hace 10.000 años y la actualidad (Figura 7). Durante este periodo, denominado Tardiglaciar, se produce la fusión de los hielos que cubrían amplias superficies terrestres y marinas. El dominio de los hielos se reduce, quedando a mediados del Holoceno restringido a las cumbres de mayor altitud. La mejora de las condiciones climáticas, determinan la expansión del bosque. Las formaciones arbóreas van ocupando cada vez más territorio hasta alcanzar su máxima extensión a mediados del Holoceno, coincidiendo con el óptimo climático holoceno. En el NW de la Península Ibérica las formaciones caducifolias de angiospermas se hacen dominantes en una gran parte de las áreas más oceánicas de la fachada cantábro-atlántica, mientras que los territorios interiores aparecen dominados por pinares o por mosaicos de pinares, bosques de angiospermas perennifolias y bosques caducifolios.
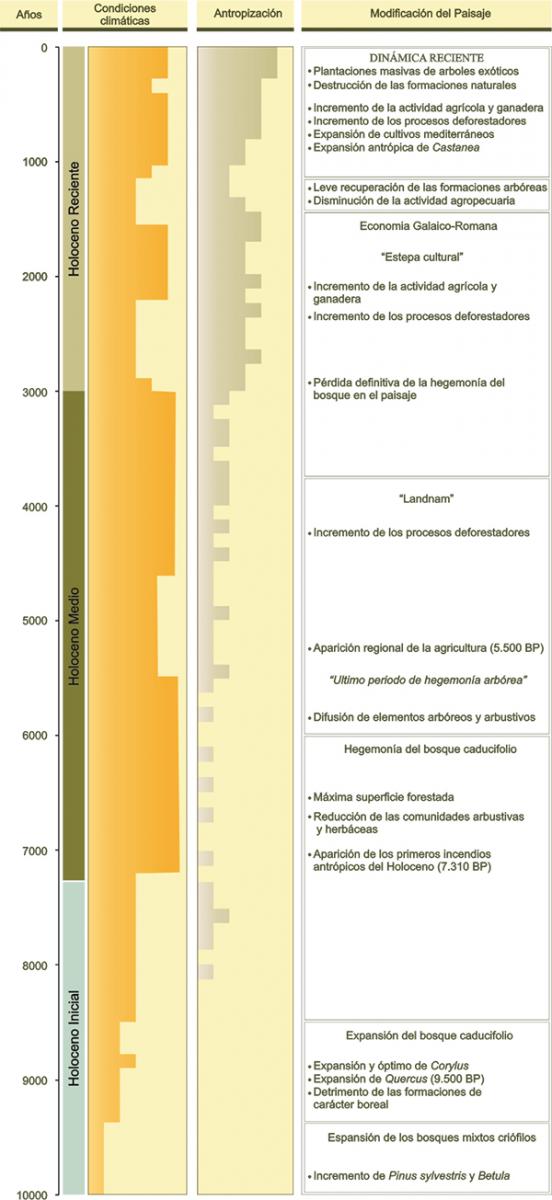
Figura 7.- Periodización ambiental del Holoceno en Galicia.
Durante las fases de deglaciación Tardiglaciar y el inicio del actual interglaciar la escasa presencia de Taxus en los diagramas del NW de la Península Ibérica atestigua la persistencia de la especie en el territorio durante el último ciclo Glaciar-Interglaciar, ocupando localidades muy distintas a las actuales, ya que estas últimas en su mayoría mostraban durante el Pleistoceno condiciones incompatibles con el desarrollo de la especie. Durante este periodo de deshielo las secuencias de Pena Vella en las sierras septentrionales de Galicia (Ramil-Rego, 1992), La Roya (Allen et al., 1986), en Sanabria y Quintanar de la Sierra (Peñalba, 1989) en Burgos, registran la presencia de Taxus (Tabla 1, Figura 8).
Dinámica de Taxus durante las fases iniciales del Holoceno
El tejo incrementa en el transcurso del Holoceno su distribución territorial, progresando en altitud en las distintas regiones del Norte de la Península Ibérica (Tabla 1 y Figura 6). Durante las fases de expansión y dominio del bosque que marcan el inicio del Holoceno, aumenta el número de registros de Taxus en el NW ibérico. De esta forma, a lo largo de la cornisa cantábrica se registran diversas presencias, tanto de granos de polen en secuencias polínicas, como de carbones en yacimientos arqueológicos (Tablas 1 y 2). En ambos casos, los datos disponibles se refieren a presencias puntuales que se corresponderían con la presencia de tejos, como individuos aislados incluidos dentro de las masas arbóreas dominantes, no detectándose signos de la presencia de tejedas.
| Depósito | Tr | HI | HM | HR | Referencia |
| Pena Vella | Ramil Rego, 1992 | ||||
| Pozo do Carballal | Muñoz Sobrino, 2001 | ||||
| La Roya | Allen et al., 1996 | ||||
| Cueto de Avellanosa | Núñez de la Fuente, 2018 | ||||
| Zalama | Pérez Díaz et al., 2016 | ||||
| Los Tornos | Peñalba, 1989 | ||||
| Los Tornos 3-5 | Peñalba, 1989 | ||||
| Quintanar de la Sierra | Peñalba, 1989 | ||||
| Belate | Peñalba, 1989 | ||||
| Atxurri | Peñalba, 1989 | ||||
| Saldropo 1 | Peñalba, 1989 | ||||
| Inurritza 1 | Peñalba, 1989 | ||||
| Fuente del Vaquero | Pérez-Díaz, 2012 |
Tabla 1.- Secuencias polínicas del NW ibérico en las que se ha identificado la presencia polen de Taxus (Tr: Tardiglaciar, HI: Holoceno Inicial, HM: Holoceno Medio, HR: Holoceno reciente).
| Depósito | HI | HM | HR | Referencia |
| Atxoste | Ruíz Alonso, 2003-2007 | |||
| Buruntza | Olaetxea, 1997 | |||
| Ondarre | Mujika et al., 2013 | |||
| Mulisko Gaina | Peñalba, 1987 | |||
| Aizpea | Zapata, 2001 | |||
| Peña Larga | Zapata, 2001 | |||
| Arrubi | Ruiz-Alonso, 2003-2007 | |||
| Mendandia | Iriarte, 2006 | |||
| Peña Parda | Ruiz-Alonso & Zapata, 2003 | |||
| El Mirón | Zapata, 2012 | |||
| Mazaculos | Uzquiano, 1995 | |||
| Aramo II | Beato et al., 2019 | |||
| Peña Oviedo | Carrión Marco, 2005 | |||
| Pala da Vella | Carrión Marco, 2003 |
Tabla 2.- Depósitos del NW ibérico en los que se ha identificado la presencia de carbones de Taxus (HI: Holoceno Inicial, HM: Holoceno Medio, HR: Holoceno reciente).
Durante las fases iniciales del Holoceno las perturbaciones humanas son reducidas y concentradas en determinadas áreas geográficas, para intensificarse y generalizarse posteriormente en el Holoceno final, especialmente en territorios como los de la Europa occidental, donde se producirá una fuerte reducción y aislamiento de los ecosistemas naturales frente a los antropoecosistemas.
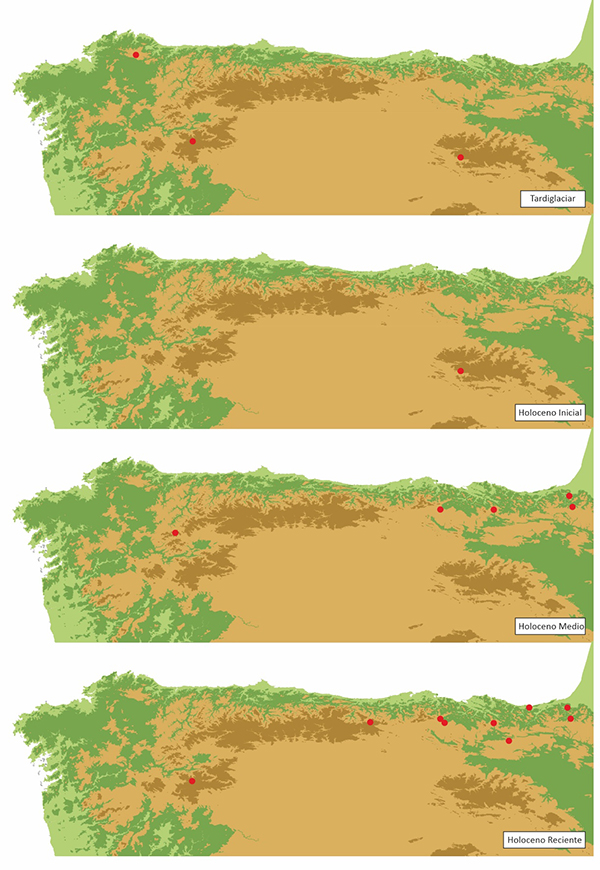
Figura 8.- Secuencias polínicas con presencia de polen de Taxus durante el Tardiglaciar y el Holoceno en el NW de la Península Ibérica.
Dinámica de Taxus durante el último tercio del Holoceno
Durante el último tercio del Holoceno las secuencias polínicas registran la aparición de polen de Taxus en algunas secuencias, mientras que en otras se pierde su registro (Figura 8). El registro de una mayor cantidad de localidades con polen de Taxus, con respecto a etapas anteriores del Holoceno, se relacionaría con la reducción de la lluvia polínica de las formaciones arbóreas dominantes, que enmascararían en muchas secuencias el polen de elementos arbóreos presentes de forma aislada entre los bosques mayoritarios. Con la reducción de la importancia paisajística del bosque, estos elementos minoritarios serían detectados con mayor facilidad en las secuencias.
La adopción de los sistemas de producción agrícola y ganadera provocará cambios fundamentales en la configuración del paisaje en toda Europa. En el Norte de la Península Ibérica este proceso se inicia hace 5.500 años cuando se registran los primeros registros de plantas cultivadas. Coincidiendo con el final de la Edad del Bronce y los inicios de la Edad del Hierro, las pequeñas aldeas de agricultores y ganaderos incrementan su producción y su huella sobre el paisaje. De modo que hace 2.000 años en las áreas más intensamente pobladas, los bosques y la vegetación natural se ve desplazada por la configuración de agrosistemas sustentados por la fuerza de las personas y de los animales domésticos.
El tejo se hace ahora escaso en el paisaje, y al igual que otras especies arbóreas queda refugiado en los enclaves montañosos donde la presión agrícola y ganadera es menos importantes. Los paisajes abiertos configurados por agrosistemas se mantienen y refuerzan durante la Edad Media y la Edad Moderna, incrementando el confinamiento de las áreas boscosas a las zonas montañosas. En muchos enclaves, especialmente de montaña, el tejo adquiere en estas épocas un protagonismo especial vinculado a su uso como recurso medicinal, simbólico y espiritual, integrándose en los espacios domésticos y religiosos. Donde permanecen como testigos vivos de un complejo y remoto pasado, mientras las formaciones nativas se desvanecen y finalmente desaparecen.
Las formaciones y bosques de tejo, son un componente básico de la biodiversidad en el territorio europeo, forjado a lo largo de miles de años como resultado de los grandes cambios biogeográficos y climáticos. Bosques y formaciones que han mantenido y todavía mantienen en muchas áreas, como ocurre en la Región Atlántica de la Península Ibérica, una intrincada relación con las comunidades locales que se han beneficiado de los servicios ecosistémicos que reportan.
Bibliografía
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Beato Bergua, S., Poblete Piedrabuena, M.A., Cunill Artigas, R. (2019). Taxus baccata en la Sierra del Aramo (Macizo Central Asturiano). Boletín de la Asociación de Geógrafos Españoles, 81, 2772, 1–30.
Camuera, J., Jiménez-Moreno, G., Ramos-Román, M.J., García-Alix, A., Toney, J.L., Anderson, R.S., Jiménez-Espejo, F., Bright, J., Webster, C., Yanes, Y., Carrión, J.S. (2019). Vegetation and climate changes during the last two glacial-interglacial cycles in the western Mediterranean: A new long pollen record from Padul (southern Iberian Peninsula),Quaternary Science Reviews, 205: 86-105.
Carrión Marco, Y. (2003). El paisaje vegetal prehistórico de Pala da Vella: primeros resultados antracológicos. In: V. Oliveira Jorge. Contributos das ciencias e das tecnologias para a arqueologia da Península Ibérica: 21-32. UTAD Vila Real, Portugal
Carrión Marco, Y. (2005). El impacto de la economía productora en el paisaje vegetal de Peña Oviedo (Camaleño, Cantabria). In: R. Ontañón Peredo, C. García-Moncó Piñeiro, P. Arias Cabal. Actas del III Congreso del Neolítico en la Península Ibérica: 35-44. Santander.
Carrión, J. S., Scott, L., Arribas, A., Fuentes, N., Gil‐Romera, G. and Montoya, E. (2006). Pleistocene landscapes in central Iberia inferred from pollen analysis of hyena coprolites. Journal of Quaternary Science, 22: 191–202.
Coxon, P., McCarron, S., Mitchell, F. (Eds) (2017). Advances in Irish Quaternary Studies. Atlantis Advances in Quaternary Science. Atlantis Press, Paris. 316pp.
Deforce, K. & Bastiaens, J. (2007). The Holocene history of Taxus baccata (yew) in Belgium and neighbouring regions. Belgian Journal of Botany, 140(2): 222-237.
Florin, R. (1958). On Jurassic taxads and conifers from North-Western Europe and Eastern Greenland. Acta Horti Bergiani Series, 17: 257-402.
Godwin H. (1975). The history of the British Flora. A factual basis for phytogeography. Cambridge University Press, Cambridge. 383 pp.
Hao, D.-C., Huang, B., Yang, L. (2008). Phylogenetic Relationships of the Genus Taxus Inferred from Chloroplast Intergenic Spacer and Nuclear Coding DNA. Biological & pharmaceutical bulletin, 31(2): 260-265.
Hartzell, J.R. (1991). The Yew Tree: A Thousand Whispers. Hulogosi Communications, Oregon, 319 pp.
Heim J. (1970). Les relations entre les spectres polliniques récents et la végétation actuelle en Europe occidentale. Ph.D. thesis, Université catholique de Louvain, Louvain-la-Neuve, Belgium.
Iriarte, M.J. (2006). El entorno vegetal del abrigo de Mendandia y su depósito arqueológico: Análisis palinológico. In: Alday A. (Dir.). El legado arqueologico de Mendandia. Los modos de vida de los últimos cazadores en la Prehistoria de Treviño: 405-418. Junta de Castilla y León. Burgos.
Jessen, K., Andersen, S.Th., Farrington, A. (1959). The interglacial deposit near Gort, Co. Galway, Ireland. Proceedings of the Royal Irish Academy B, 60: 1-77.
Kasse K. (1988). Early-Pleistocene tidal and fluviatile environments in the southern Netherlands and northern Belgium. Ph.D. thesis, Vrije Universiteit, Amsterdam, The Netherlands.
Kelly, M.R. (1964) The Middle Pleistocene of North Birmingham. Philos. T. Roy. Soc. B, 247: 533-592.
Kunzmann L., Mai D.H. (2005) Die Koniferen der Mastixioideen-Flora von Wiesa bei Kamenz (Sachsen, Miozän) unter besonderer Berücksichtigung der Nadelblätter. Palaeontographica B 272(1–6): 67–135.
Leroy, S.A.G. (2008): Vegetation cycles in a disturbed sequence around the Cobb-Mountain subchron in Catalonia (Spain). Journal of Paleolimnology, 40(3), 851-868
Li, J., Davis, C.C., Del Tredici, P., Donoghue, M.J. (2001). Phylogeny and biogeography of Taxus (Taxaceae) inferred from sequences of the internal transcribed spacer region of nuclear ribosomal DNA. Harv. Pap. Bot. ,6:267–274.
Mujika-Alustiza, J.A., Agirre-García, J., Edeso-Fito, J.M., Lopetegi-Galarraga, A., Pérez-Díaz, S., Ruiz-Alonso, M., Tarriño-Vinagre, A., Yusta-Arnal, I. (2013). La continuidad de la actividad pastoril durante la época romana en la zona de Argarbi (sierra de Aralar - Gipuzkoa). Kobie Serie PaleoantroPología, 32: 217-258
Muñoz Sobrino, C. (2001). Cambio climático y dinámica del paisaje en las montañas del noroeste de la Península Ibérica. Tesis Doctoral. Escuela Politécnica Superior. Universidad de Santiago de Compostela.
Nathorst A.G. (1908). Palaeobotanische Mitteillungen.7. Über Palyssya, Stachyotaxus und Paleotaxus. Ksvenka Vetensk Akad Handl 8: 1–16.
Nuñez de la Fuente, S. (2018). Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje atlántico montañoso: la región cantábrica durante el Holoceno. Tesis doctoral. Universidad de Cantabria.
Peñalba Garmendia, M.C. (1989). Dynamique de Végétation Tardiglaciaire et Holocene Du Centre-Nord de l’Espagne d’après l’analyse pollinique. These. Université d’Aix, Marseille III.
Pérez-Díaz, S. (2012). El paisaje vegetal durante la Prehistoria Reciente en la vertiente mediterránea de Euskal Herria— Tesis doctoral. Euskal Herriko Univ., Vitoria-Gastei.
Perez-Díaz, S., López-Sáez, J. A., Pontevedra-Pombal, X., Souto-Souto, M. & Galop, D. (2016). 8000 years of vegetation history in the northern Iberian Peninsula inferred from the palaeoenvironmental study of the Zalama ombrotrophic bog (Basque-Cantabrian Mountains, Spain). Boreas, Vol. 45, pp. 658–672.
Ramil Rego P, Gómez-Orellana L, Muñoz Sobrino C, Tereso JP (2011). Dinâmica natural e transformação antrópica das florestas do noroeste ibérico. In Tereso JP, Honrado JP, Pinto AT, Rego FC (Eds.) Florestas do Norte de Portugal: História, Ecologia e Desafios de Gestão. InBio - Rede de Investigação em Biodiversidade e Biologia Evolutiva. Porto. ISBN: 978-989-97418-1-2. Pp 14-54.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ruiz-Alonso, M. & Zapata, L. (2003). Análisis antracológico del yacimiento arqueológico de Peña Parda. Cuad. Arq. Univ. Navarra, 11: 217-252.
Ruiz-Alonso, M. (2003/2007). Madera carbonizada en los fondos de cabaña de Arrubi y Esnaurreta (Sierra de Aralar, Gipuzkoa): vegetación y recursos forestales en la Edad Media. Kobie (Serie Paleoantr.), 27: 131-150.
Uzquiano, P. (1995). L’evolution de la vegetation a l’Holocene inicial dans le nord de l’Espagne a partir de l’étude anthracologique de trois sites archeologique. Quaternaire 6 (2): 77-83.
Van der Knaap, W.O., van Leeuwen, J.F.N. (1997). Late Glacial and early Holocene vegetation succession, altitudinal vegetation zonation, and climatic change in the Serra da Estrela, Portugal. Review of Palaeobotany and Palynology, 97:3-4: 239-285.
Watts, W.A. (1985) Quaternary vegetation cycles.In: Edwards, K.J. & Warren, W.P. (eds.): The Quaternary history of Ireland: 154-185. Academic Press, London.
West, R.G. (1962) A note on Taxus pollen in the Hoxnian Interglacial. New Phytologist, 61: 189-190.
Zapata, L. (2012). Holocene wood charcoal from El Mirón cave. Vegetation and wood use — In: Straus, L.G. & González Morales, M.R. (Eds.). El Miron cave. Cantabrian Spain. The site and its Holocene archaeological record: 174-196. Univ. New Mexico, Alburquerque.